Orchestrating the Mouse Gastrula: An Integrated View of BMP, Nodal, and FGF Signaling Pathways
Gastrulation is a pivotal event in mammalian development, establishing the three primary germ layers through the precise coordination of key signaling pathways.
Orchestrating the Mouse Gastrula: An Integrated View of BMP, Nodal, and FGF Signaling Pathways
Abstract
Gastrulation is a pivotal event in mammalian development, establishing the three primary germ layers through the precise coordination of key signaling pathways. This article provides a comprehensive synthesis for researchers and drug development professionals on the roles of BMP, Nodal, and FGF signaling during mouse gastrulation. It explores the foundational principles of how these pathways direct lineage specification and embryonic patterning, examines advanced methodological approaches for their study—including embryo models and single-cell technologies—and addresses critical challenges in pathway modulation and experimental optimization. Furthermore, it discusses validation and comparative strategies that leverage epigenetic, transcriptomic, and cross-species analyses to confirm pathway functions and resolve model discrepancies, offering an integrated resource for developmental biology and regenerative medicine applications.
Core Principles: How BMP, Nodal, and FGF Direct Germ Layer Formation
Gastrulation is a pivotal period in early mammalian development during which the pluripotent epiblast is transformed into the three definitive germ layers—ectoderm, mesoderm, and endoderm—that form the foundation for all fetal tissues and organs [1] [2]. In the mouse, this process occurs mainly between embryonic day (E) 6.25 and E9.5 and is characterized by extensive cellular reorganization, including the formation of the primitive streak, a key morphological structure that serves as a portal for the emergence of mesodermal and endodermal progenitors [1]. The precise execution of germ layer specification is orchestrated by a dynamic and integrated network of signaling pathways, primarily BMP, Nodal, Wnt, and Fgf, which establish concentration gradients and temporal sequences that pattern the embryo [3] [4] [1]. Understanding the mechanisms of mouse gastrulation provides fundamental insights into embryogenesis and has profound implications for regenerative medicine, disease modeling, and stem cell engineering.
The Establishment of the Embryonic Axis
The process of gastrulation is predicated on the prior establishment of the Anterior-Posterior (AP) axis, which breaks the radial symmetry of the early embryo and defines the site of primitive streak formation [1].
From Proximal-Distal to Anterior-Posterior Polarity
Before gastrulation, the mouse conceptus is radially symmetrical around the proximal-distal (PD) axis. The transformation into a bilaterally symmetrical embryo with a defined AP axis involves a coordinated sequence of events [1]:
- Specification of the Distal Visceral Endoderm (DVE): A subset of distal visceral endoderm cells is specified at the distal pole of the conceptus in response to low levels of Nodal signaling and the inhibition of BMP4 from the extra-embryonic ectoderm [1].
- Migration to form the Anterior Visceral Endoderm (AVE): The DVE migrates proximally to the future anterior side, forming the AVE. This migration direction determines the anterior of the embryo [1].
- Restriction of Primitive Streak Formation: The AVE secretes inhibitors such as CER1, LEFTY1 (Nodal antagonists), and DKK1 (Wnt antagonist), which restrict the signaling domains that induce the primitive streak to the posterior pole [1].
Table: Key Signaling Centers in Axis Formation
| Signaling Center | Location | Key Signals Expressed | Primary Function |
|---|---|---|---|
| Extra-Embryonic Ectoderm (ExE) | Proximal | BMP4, Nodal convertases (FURIN/PACE4) | Inhibits DVE formation; activates Nodal to create PD gradient [1] |
| Distal Visceral Endoderm (DVE) | Distal pole | CER1, LEFTY1 (Nodal inhibitors) | Initiates symmetry breaking; begins inhibition of posterior signals [1] |
| Anterior Visceral Endoderm (AVE) | Anterior | CER1, LEFTY1, DKK1 (Wnt inhibitor) | Restricts primitive streak formation to the posterior by inhibiting Nodal and Wnt [1] |
| Posterior Epiblast | Posterior | WNT3a, NODAL | Induces and positions the primitive streak [1] |
Signaling Gradients in Axis Patterning
The establishment of the AP axis is a classic example of morphogen gradient-driven patterning. The proximal epiblast produces pro-NODAL, which is converted to its active form by convertases in the extra-embryonic ectoderm, creating a Nodal signaling gradient from proximal to distal [1]. The migration of the AVE then transforms this radial gradient into an anterior-posterior gradient, with high Nodal and Wnt activity posteriorly and low activity anteriorly [1]. In zebrafish, it has been shown that opposing gradients of Nodal and BMP are sufficient to induce a complete embryonic axis, and the ratio of pSmad2 (Nodal effector) to pSmad5 (BMP effector) activity is a critical determinant of cell fate [5].
Core Signaling Pathways in Gastrulation
The following diagram illustrates the core signaling network and its functional outcomes during mouse gastrulation.
BMP Signaling
- Role: BMP signaling is essential for inducing the primitive streak and is particularly critical for specifying the extra-embryonic mesoderm lineages, which contribute to structures like the allantois and yolk sac mesenchyme [1] [6].
- Source: The primary source of BMP (e.g., BMP4) in the early mouse embryo is the extra-embryonic ectoderm [1].
- Mechanism: BMPs signal through receptor-activated Smad proteins (e.g., Smad1/5/8). In zebrafish, the gradient of phosphorylated Smad5 (pSmad5) provides a broad posteriorizing signal, and its ratio to pSmad2 helps determine cell fate [5].
Nodal Signaling
- Role: Nodal, a TGF-β family ligand, is a master regulator of primitive streak formation, mesendoderm specification, and the establishment of left-right asymmetry [4] [1].
- Source and Gradient: A proximal-to-distal gradient of active Nodal is established before gastrulation, which is later refined into an anterior-posterior gradient by the action of the AVE [1].
- Mechanism: Nodal signals through Smad2/3. The duration of Nodal signaling is as critical as its concentration in gradient interpretation and cell fate specification [4]. Nodal and BMP signaling effectors, Smad2 and Smad5, exhibit mutual antagonism, providing a mechanism for cells to integrate and discriminate between these overlapping signals [5].
Wnt Signaling
- Role: Wnt signaling (e.g., via WNT3a) is required for the induction and posterior maintenance of the primitive streak [1]. It is also crucial for the formation and maintenance of neuromesodermal progenitors (NMPs), a bipotent population that gives rise to the spinal cord and paraxial mesoderm [1] [7].
- Regulation: The anterior extent of the Wnt signaling domain is restricted by the Wnt inhibitor DKK1, secreted by the AVE [1].
Fgf Signaling
- Role: The FGF/ERK pathway is a key regulator of cell potency and lineage decisions in the epiblast [6]. It promotes the acquisition of a posterior mesodermal fate and is involved in the epithelial-to-mesenchymal transition (EMT) required for ingression through the primitive streak [1] [6].
- Gradient and Epigenetics: The FGF/ERK pathway displays a gradient of activity within the epiblast, with higher activity in the anterior. This activity influences the epigenetic state of epiblast cells, predetermining their response to differentiation signals by modulating DNA methylation and chromatin accessibility [6].
Table: Signaling Pathway Roles in Germ Layer Specification
| Signaling Pathway | Primary Role in Gastrulation | Key Germ Layer Contributions | Major Inhibitors |
|---|---|---|---|
| BMP | Induces primitive streak; specifies extra-embryonic mesoderm [1] [6] | Mesoderm (especially extra-embryonic) [6] | CHORDIN, NOGGIN |
| Nodal | Induces primitive streak; specifies mesendoderm [4] [1] | Mesoderm, Endoderm [4] | LEFTY1, CER1 (from AVE) [1] |
| Wnt | Maintains primitive streak; specifies NMPs [1] [7] | Mesoderm (posterior), Ectoderm (posterior neural) via NMPs [7] | DKK1 (from AVE) [1] |
| Fgf | Regulates EMT; maintains primed state; patterns mesoderm [1] [6] | Mesoderm, influences Ectoderm/Endoderm via epigenetic priming [6] | ERK inhibitors (e.g., PD03) [6] |
Key Experimental Models and Methodologies
The study of mouse gastrulation employs a combination of in vivo and in vitro models, each offering unique advantages for dissecting complex developmental processes.
In Vivo Mouse Embryo Studies
The foundational knowledge of gastrulation has been built on decades of genetic experimentation in mouse embryos [3].
- Key Techniques:
- Genetic Fate Mapping: Historically used to map the developmental potential of different regions of the epiblast, establishing the basic fate map of the mouse embryo [1].
- Mutant Analysis: Studying embryos with targeted mutations in genes of signaling pathways (e.g., Nodal, BMP receptors) to determine their functional requirements [1].
- Advanced Imaging and Single-Cell 'Omics: Modern techniques like live imaging, single-cell RNA sequencing (scRNA-seq), and spatial transcriptomics allow for the capture of dynamic processes and high-resolution molecular profiling of individual cells across the entire embryo [1] [6].
In Vitro Embryo Models
Recent years have seen a surge in the use of embryo models to overcome the technical challenges of studying early mammalian development in utero [3] [1].
- Gastruloids: These 3D aggregates of mouse embryonic stem (ES) cells self-organize and recapitulate key aspects of gastrulation, including the expression of axial markers and the emergence of germ layer derivatives [3]. They provide a powerful, high-throughput platform for real-time visualization and manipulation of signaling dynamics.
- EpiSC Differentiation: Mouse epiblast stem cells (EpiSCs), derived from the post-implantation epiblast, are a 2D model for studying the primed state of pluripotency and lineage commitment [6]. They can be differentiated toward various germ layer fates in response to defined signaling cues.
The following workflow outlines a typical experimental approach integrating these models to study signaling in gastrulation.
Detailed Protocol: Investigating Epiblast Priming using EpiSCs
This protocol, based on the work of [6], leverages EpiSCs to investigate how intrinsic epigenetic states prime cells for specific lineage fates.
- Cell Line: Mouse EpiSCs (e.g., EpiSC line).
- Culture Maintenance: Maintain EpiSCs in a primed state using culture medium supplemented with FGF2 (Fgf/ERK signaling activator) and Activin A (TGF-β/Smad2/3 signaling activator) [6].
- Separation of Regionalized Epiblast Populations:
- Dissociate EpiSCs into a single-cell suspension.
- Perform Fluorescence-Activated Cell Sorting (FACS) using an antibody against the surface marker CLDN6. This separates cells into CLDN6High (representing anterior epiblast) and CLDN6Low (representing distal posterior epiblast) populations [6].
- Lineage Differentiation Assay:
- Seed sorted CLDN6High and CLDN6Low populations separately.
- Subject them to differentiation protocols that provide high WNT and TGF-β signaling to drive differentiation towards definitive endoderm and other germ layers [6].
- Alternatively, treat with BMP4 to assess extra-embryonic mesoderm potential [6].
- Downstream Analysis:
- Flow Cytometry: Quantify phosphorylated ERK (pERK) levels to correlate FGF/ERK signaling activity with CLDN6 status [6].
- Single-Cell RNA-seq: Profile the transcriptomes of differentiating cells to assess lineage bias and identify differentially expressed genes.
- Epigenetic Analysis: Perform assays for chromatin accessibility (ATAC-seq) and DNA methylation (whole-genome bisulfite sequencing) on the sorted populations to identify priming epigenetic signatures [6].
The Scientist's Toolkit: Key Research Reagents
Table: Essential Reagents for Studying Mouse Gastrulation
| Reagent / Tool | Category | Example(s) | Primary Function in Research |
|---|---|---|---|
| Signaling Agonists | Small Molecules/Proteins | Recombinant BMP4, Nodal, FGF2, Wnt3a, Activin A (mimics Nodal) | Activate specific signaling pathways to probe their function in differentiation protocols [6] [7] |
| Signaling Antagonists | Small Molecules/Proteins | LDN193189 (BMP inhibitor), SB431542 (Nodal/TGF-β inhibitor), PD03 (ERK inhibitor), DKK1 (Wnt inhibitor) | Inhibit specific signaling pathways to assess necessity and define signaling windows [6] |
| Cell Surface Markers | Antibodies | Anti-CLDN6, Anti-E-CADHERIN | Isate distinct regional epiblast populations (e.g., by FACS) for functional analysis [6] |
| Lineage Reporters | Genetically Modified Cells | Sox1-GFP (neuroectoderm), Bra-GFP (mesoderm), Sox17-GFP (endoderm) | Visualize and isolate specific germ layer derivatives in real time during differentiation [1] |
| Epigenetic Assays | Kits/Reagents | ATAC-seq kit, Bisulfite Conversion kit | Profile chromatin accessibility and DNA methylation states to investigate lineage priming [6] |
Mouse gastrulation represents a paradigm of how coordinated signaling input is translated into organized tissue output. The integrated activities of the BMP, Nodal, Wnt, and Fgf pathways establish a dynamic signaling landscape that is interpreted by epiblast cells whose epigenetic state and spatial position predetermine their response, leading to the specification of the primary germ layers [3] [1] [6]. The ongoing development of sophisticated in vitro models like gastruloids, coupled with high-resolution omics technologies and computational modeling, promises to further deconvolute the exquisite complexity of this process [3] [1]. This deeper understanding is not only fundamental to developmental biology but also critically informs efforts to direct the differentiation of stem cells for regenerative medicine and model human developmental disorders.
Gastrulation is a fundamental process in mammalian embryonic development during which the pluripotent epiblast self-organizes into the three primary germ layers—the endoderm, mesoderm, and ectoderm—that will form the entire embryo. Decades of research, primarily in the mouse model, have established that a conserved signaling cascade involving the Bone Morphogenetic Protein (BMP), WNT, and NODAL pathways is indispensable for initiating gastrulation and patterning the primitive streak [8] [3]. This review focuses on the critical role of the BMP pathway, framing its function within the broader, dynamic signaling network that orchestrates cell fate decisions in the early mouse embryo. BMP signaling does not operate in isolation; it is embedded in a complex regulatory circuitry with FGF and other pathways, which collectively ensure the reproducible specification and spatial organization of mesodermal precursor populations [9]. Understanding the quantitative principles and mechanistic interactions of this network is crucial for researchers and drug development professionals aiming to manipulate cell fate in regenerative medicine or model developmental disorders.
The Core Signaling Network and Its Regulatory Logic
The BMP-WNT-NODAL Signaling Hierarchy
In the mouse embryo, gastrulation is initiated on the posterior side, where a signaling center becomes active. The prevailing model, supported by both in vivo and in vitro studies, posits a hierarchical cascade: BMP signaling activates WNT signaling, which in turn activates NODAL signaling [8]. This cascade is integral to the formation of the primitive streak. In vitro models of human gastrulation have quantified the dynamics of this process, revealing that BMP signaling initiates waves of WNT and NODAL signaling activity that propagate toward the center of a cell colony at a constant rate [8]. This wave-like behavior is inconsistent with a simple, stable reaction-diffusion gradient (Turing system). Instead, the final signaling state tends toward homogeneity, with spatial differences arising primarily from boundary effects and the dynamic history of signal exposure [8] [10].
The following diagram illustrates the core signaling hierarchy and key regulatory interactions that govern primitive streak patterning:
The Antagonistic Role of FGF Signaling
While BMP, WNT, and Nodal form the core inductive cascade, FGF signaling plays a pivotal and often antagonistic role in mesoderm patterning. Single-cell resolution studies in differentiating stem cell populations have identified opposing functions for BMP and FGF [9]. FGF stimulation boosts the expression of endogenous Fgf genes while simultaneously repressing Bmp ligand genes. This positive autoregulation of FGF, coupled with its repression of the BMP pathway, creates a binary switch mechanism. This regulatory logic contributes to a "community effect," ensuring the specification of coherent cohorts of cells with the same identity, thereby helping to segregate BMP-dependent proximal mesoderm cell types from FGF-dependent distal mesoderm cell types [9].
Quantitative BMP Signaling and Fate Determination
Concentration-Dependent Fate Patterning
The concentration of BMP ligand is a primary determinant of cell fate. Studies using epiblast stem cell (EpiSC) differentiation protocols have systematically mapped how BMP4 concentration guides cells toward distinct mesodermal subtypes, consistent with its proposed gradient-based activity in the embryo [9].
Table 1: BMP4 Concentration-Dependent Cell Fate Specification in EpiSC Differentiation
| BMP4 Concentration | Key Marker Expression | Inferred Mesoderm Subtype | Embryonic Region |
|---|---|---|---|
| Low (∼1-4 ng/mL) | Peak Foxa2, Shh, Tbx6, Msgn1 |
Axial & Paraxial Mesoderm | Distal (Posterior) |
| Medium (∼4-8 ng/mL) | Peak T/Bra, Tbx6 |
General & Distal Mesoderm | Distal |
| High (≥16 ng/mL) | Peak Hand1, Gata6 |
Extraembryonic & Lateral Plate Mesoderm | Proximal |
Temporal Dynamics and Signal Integration
A critical advance in understanding BMP signaling has been the recognition that signaling duration, not just amplitude, controls cell fate decisions. Live-cell imaging in micropatterned human pluripotent stem cell (hPSC) models has revealed that the time-integral of BMP signaling is the primary determinant of cell fate [10].
In this model, the level and duration of BMP signaling are interchangeable parameters; a lower signal level over a longer duration can induce the same fate as a higher signal level over a shorter duration, provided the time-integral of signaling is equivalent [10]. Mechanistically, this integration is mediated by the transcription factor SOX2, which represses differentiation genes. The level of SOX2 decreases in proportion to the time-integral of BMP signaling, thereby permitting the activation of fate-specific gene programs [10].
Table 2: BMP Signaling Dynamics and Correlated Cell Fates in a 2D Gastruloid Model
| Signaling History Cluster | BMP Signaling Dynamics | Nodal Signaling Activity | Resultant Cell Fate |
|---|---|---|---|
| Sustained High | High signaling maintained throughout | Low | Amnion-like / Extraembryonic |
| Transient | High initial signaling, then low | Low | Pluripotent Epiblast-like |
| Biphasic | High initial signaling, then low, then high again (Nodal wave) | High | Primitive Streak-like / Mesoderm |
The following diagram visualizes this fate decision mechanism based on the time-integrated BMP signal:
Experimental Models and Methodologies
Key In Vitro Models for Studying Gastrulation
To overcome the challenges of studying signaling dynamics in vivo, several powerful in vitro models have been developed:
- 2D Micropatterned hPSC Gastruloids: This high-throughput system involves confining hPSCs to small, circular micropatterns and stimulating them with BMP4. The cells self-organize into concentric rings of distinct fates: an outer ring of CDX2+ extraembryonic cells, a middle ring of BRA+ mesodermal cells, and a central disc of NANOG+/SOX2+ pluripotent cells [8] [10]. This model is ideal for quantitative imaging and perturbation studies.
- EpiSC Differentiation Protocols: Mouse or human Epiblast Stem Cells (EpiSCs) can be differentiated in a more homogeneous culture system. Cells are typically transitioned from a pluripotency-maintaining medium (e.g., containing ActivinA and FGF2) to a differentiation medium containing a WNT agonist (e.g., CHIR99021) and titrated amounts of BMP4 and FGF to assess concentration-dependent effects on mesoderm specification [9].
Protocol: Analyzing BMP-Mediated Patterning in 2D Gastruloids
1. Cell Seeding and Micropatterning:
- Use a defined medium such as mTeSR1.
- Seed hPSCs onto micropatterned substrate discs (e.g., 200-500 µm diameter) coated with a suitable adhesion protein (e.g., Matrigel, Laminin-521).
- Allow cells to attach and form a confluent colony overnight.
2. BMP4 Induction:
- Initiate differentiation by adding a defined concentration of BMP4 (e.g., 10-50 ng/mL) to the medium.
- Maintain the culture for 48-72 hours for pattern formation.
3. Live-Cell Imaging of Signaling Dynamics:
- Use hPSC reporter lines expressing fluorescently tagged SMAD proteins (e.g., GFP::SMAD4 for BMP/Nodal activity, RFP::SMAD1 for specific BMP activity).
- Image nuclei over 48-72 hours using live-cell microscopy, tracking SMAD nuclear/cytoplasmic intensity ratios as a proxy for pathway activity.
4. Endpoint Immunostaining and Analysis:
- Fix gastruloids at the desired time point.
- Perform iterative immunofluorescence staining for key fate markers:
- CDX2 for extraembryonic fate.
- BRA (Brachyury) for mesodermal fate.
- SOX17 for endodermal fate.
- NANOG/SOX2 for pluripotent epiblast.
- Correlate the final fate of each cell with its previously recorded signaling history to establish predictive relationships [10].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Investigating BMP Signaling in Gastrulation Models
| Reagent / Tool | Function / Target | Example Use Case | Citation |
|---|---|---|---|
| Recombinant BMP4 | Ligand; activates BMP receptor complex | Induce differentiation in micropatterned gastruloids | [8] [9] |
| K02288 | Inhibitor of BMP type I receptors (ACVR1, BMPR1A) | Chemically inhibit BMP-SMAD signaling to test necessity | [11] |
| IWP2 | Inhibitor of WNT secretion | Disrupt the BMP-WNT-NODAL hierarchy; blocks Nodal wave | [8] |
| GFP::SMAD4 / RFP::SMAD1 Reporter Cell Lines | Live-cell imaging of BMP/Nodal (SMAD4) or specific BMP (SMAD1) signaling | Quantify spatiotemporal signaling dynamics | [10] |
| CRISPR-Cas9 Gene Knockout | Genetic ablation of signaling components (e.g., NODAL, ACVR1) | Establish requirement of specific genes for fate decisions | [8] [11] |
| shRNA Knockdown | Transient gene silencing (e.g., of ACVR1) | Validate hits from genetic screens | [11] |
BMP signaling is a cornerstone of the regulatory network that controls primitive streak patterning and cell fate induction during mouse gastrulation. Its function is not defined by a static concentration gradient alone but is dynamically interpreted by cells through the integration of signal level and duration. This temporal control, coupled with its intricate feedback and feedforward interactions with WNT, NODAL, and antagonistic FGF pathways, ensures the robust self-organization of the early embryo. The continued use of quantitative in vitro models, single-cell technologies, and sophisticated computational approaches will be essential to fully unravel the complex logic of this developmental signaling network.
Nodal, a transforming growth factor-β (TGF-β) family member, serves as a master regulator during vertebrate embryogenesis, executing concentration-dependent functions that establish the embryonic axes, induce mesoderm and endoderm formation, and determine left-right asymmetry. This whitepaper examines the molecular mechanisms of Nodal signaling within the broader signaling network of mouse gastrulation, integrating recent advances from genetic studies and innovative stem cell-based embryo models. We detail how Nodal functions through canonical Smad2 transcription factors and FoxH1 coactivators, requires EGF-CFC coreceptors, and is antagonized by Lefty and Cerberus proteins. Emerging research reveals that Nodal operates predominantly through heterodimerization with Gdf3, a mechanism that enhances signaling potency and precision. The complex interplay between Nodal, BMP, Wnt, and FGF pathways creates a robust signaling network that orchestrates embryonic patterning through cross-regulatory interactions and community effects. Technical advances in single-cell analysis and live-imaging of signaling dynamics are refining our understanding of how signaling histories determine cell fate, offering new insights for developmental biology and regenerative medicine applications.
Nodal signaling represents one of the fundamental patterning systems in vertebrate development, directing the fate of naive embryonic cells into specialized tissues that form the foundational architecture of the organism. As a member of the TGF-β superfamily, Nodal exerts concentration-dependent effects that establish the basic body plan during gastrulation. In murine development, Nodal functions within a network of signaling pathways including BMP, Wnt, and FGF that collectively coordinate the cellular rearrangements and differentiation events that generate the three germ layers [12]. The precise spatiotemporal control of Nodal signaling activity is regulated at multiple levels—from ligand processing and secretion to receptor engagement and intracellular transduction—ensuring proper embryonic patterning. Disruption of this finely tuned system leads to severe developmental defects, underscoring its critical importance in embryogenesis. This technical guide examines the molecular machinery of Nodal signaling, its integration with other key pathways, and the experimental approaches used to decipher its functions within the context of mouse gastrulation.
Core Nodal Signaling Machinery
Molecular Components and Pathway Architecture
The Nodal signaling pathway operates through a canonical TGF-β signaling cascade with specialized components that confer its unique developmental functions. The core pathway consists of:
- Ligands: Nodal proteins are synthesized as preproproteins containing a prodomain and mature ligand domain. Following dimerization and cleavage, mature ligands initiate signaling [13].
- Receptors and Coreceptors: Nodal signals through activin type I and type II serine/threonine kinase receptors. Essential to Nodal specificity are the EGF-CFC family proteins (such as Cripto and Cryptic), which act as obligate coreceptors [14].
- Intracellular Transducers: Receptor activation leads to phosphorylation of Smad2 and Smad3 transcription factors, which then form complexes with Smad4 [14].
- Transcriptional Effectors: The Smad complexes translocate to the nucleus where they interact with coactivators, primarily FoxH1, to regulate expression of target genes including Nodal itself (autoregulation), Pitx2, and Lefty [14].
Regulatory Mechanisms and Modulators
Nodal signaling is precisely modulated at multiple levels to ensure proper patterning:
- Antagonists: Extracellular inhibitors including Lefty and Cerberus proteins antagonize Nodal signaling by preventing receptor binding [14].
- Ligand Processing: Proprotein convertases regulate the generation of mature Nodal ligands, providing an additional layer of control [14].
- Cellular Response Modulators: Intracellular factors such as Arkadia and DRAP1 enhance or suppress signaling responses, respectively [14].
Table 1: Core Components of the Nodal Signaling Pathway
| Component Type | Key Molecules | Function |
|---|---|---|
| Ligands | Nodal, Gdf3 (heterodimer) | Initiate signaling; heterodimerization enhances potency |
| Receptors | Activin receptors (Type I/II) | Serine/threonine kinase activity; phosphorylate Smads |
| Coreceptors | EGF-CFC family (Cripto) | Obligate coreceptors essential for signal transduction |
| Signal Transducers | Smad2, Smad3, Smad4 | Form transcription factor complexes |
| Transcription Factors | FoxH1 | DNA-binding coactivators for Smad complexes |
| Extracellular Antagonists | Lefty, Cerberus | Bind ligands/prevent receptor interaction |
| Feedback Regulators | Arkadia, DRAP1 | Enhance or suppress cellular responses |
Figure 1: Nodal Signaling Pathway Architecture. Nodal and Gdf3 form heterodimers that bind receptors and coreceptors, initiating intracellular Smad signaling. Target genes include Lefty, which provides negative feedback.
Heterodimerization with Gdf3: Enhanced Signaling Precision
Recent research has clarified that Nodal functions predominantly through heterodimerization with Gdf3 (also known as Gdf1 or Vg1), a related TGF-β family member. This partnership significantly enhances signaling potency and precision during embryonic patterning [13].
Mechanism of Heterodimer Formation
The heterodimerization process involves:
- Co-expression Requirement: Nodal and Gdf3 must be co-expressed in the same cells for functional signaling to occur [13].
- Intracellular Processing: Gdf3 cannot be cleaved or secreted without co-expression with Nodal. The prodomain of Gdf3 remains uncleaved unless Nodal is present [13].
- Secretion Dependency: Gdf3 secretion is entirely dependent on co-expression with Nodal, ensuring that only heterodimers are released from producing cells [13].
Functional Advantages of Heterodimerization
The Nodal-Gdf3 heterodimer provides several developmental advantages:
- Increased Potency: Heterodimers exhibit significantly enhanced signaling capability compared to Nodal homodimers, enabling more robust patterning [13].
- Temporal Control: Maternal Gdf3 persists to post-gastrulation stages, allowing it to function with zygotic Nodal in left-right patterning without requiring new Gdf3 synthesis [13].
- Spatial Precision: The co-expression requirement creates a built-in mechanism restricting signaling activity to specific domains where both ligands are produced [13].
Nodal in Embryonic Axis Formation and Mesendoderm Patterning
Role in Germ Layer Specification
During mouse gastrulation, Nodal performs concentration-dependent functions in germ layer formation:
- Endoderm Induction: High Nodal signaling promotes definitive endoderm specification [14].
- Mesoderm Patterning: Intermediate Nodal levels direct formation of various mesodermal subtypes [14] [9].
- Axial Patterning: Nodal works in concert with Wnt and BMP signals to establish the anterior-posterior axis [12].
The patterning function of Nodal is integrated with other signaling pathways through a complex transcriptional hierarchy that operates during self-organization in pluripotent cell populations [10].
Left-Right Axis Determination
Nodal plays a specialized role in establishing left-right asymmetry:
- Asymmetric Expression: Following gastrulation, Nodal exhibits asymmetric expression on the left side of the embryo [14].
- Downstream Targets: Nodal signaling activates transcription of Pitx2, which executes the developmental program for left-sided organ morphogenesis [14].
- Maternal Contribution: Maternal Gdf3 provision enables left-right patterning without requiring zygotic Gdf3 expression [13].
Table 2: Nodal Signaling Functions in Mouse Gastrulation
| Developmental Process | Nodal Function | Interacting Pathways |
|---|---|---|
| Primitive Streak Induction | Induces and patterns the primitive streak | Wnt, BMP, FGF |
| Mesendoderm Specification | Directs mesoderm and endoderm formation | Wnt (synergistic) |
| Anterior-Posterior Patterning | Establishes anterior identity | BMP (antagonistic) |
| Left-Right Asymmetry | Determines left-side identity | Gdf3 (obligate partner) |
| Self-Regulation | Maintains expression through autoregulation | FoxH1-dependent |
Signaling Pathway Integration in Mouse Gastrulation
Nodal does not function in isolation but operates within an integrated network of signaling pathways that collectively coordinate gastrulation events.
Cross-Activation and Antagonism with BMP, Wnt, and FGF
The signaling environment during gastrulation features extensive cross-talk:
- BMP and FGF Antagonism: Recent single-cell analyses reveal that FGF and BMP signaling exhibit functional antagonism during mesoderm differentiation. FGF signaling boosts expression of endogenous Fgf genes while repressing Bmp ligand genes, creating a community effect that promotes coherent domain formation [9].
- Pathway Cross-Activation: BMP signaling induces cross-activation of FGF, Nodal, and WNT pathways in mouse embryonic stem cells, creating constraints on BMP-mediated induction of the totipotent state [15].
- Transcriptional Hierarchy: In micropatterned human pluripotent stem cells, BMP, Wnt, and Nodal function in a transcriptional hierarchy during self-organized pattern formation, with Wnt and Nodal required for primitive streak-like differentiation [10].
Temporal Dynamics of Signaling Activity
Signaling pathways exhibit dynamic behavior that determines developmental outcomes:
- Signaling Histories: Single-cell tracking reveals that BMP signaling history (time-integrated activity) correlates strongly with cell fate decisions, suggesting similar temporal integration likely occurs for Nodal signaling [10].
- Dynamic Gradients: Signaling gradients change over time as the embryo develops, with cells experiencing different signaling levels throughout competence windows [10].
- Wave Patterns: Live imaging of SMAD4 signaling shows waves of activity that transition from uniform to spatially restricted patterns, with qualitative differences in signaling histories predicting distinct cell fates [10].
Figure 2: Signaling Pathway Interactions in Gastrulation. BMP, FGF, WNT, and Nodal pathways exhibit cross-activation and antagonism that collectively pattern the germ layers.
Experimental Approaches and Methodologies
Stem Cell-Based Embryo Models
Recent advances in embryo models have provided powerful platforms for studying Nodal signaling:
Gastruloid Systems: Mouse embryonic stem cell-derived gastruloids enable real-time visualization of signaling dynamics during primitive streak formation [12]. These models recapitulate key aspects of in vivo development while allowing experimental manipulation of signaling pathways.
Protocol: EpiSC Differentiation for Mesoderm Patterning Studies [9]
- Culture Conditions: Maintain epiblast stem cells (EpiSCs) in N2B27 medium containing ActivinA (to maintain pluripotency), FGF2, and the Wnt signaling inhibitor XAV939 (FAX medium) to obtain a homogeneous starting population.
- Mesoderm Induction: Trigger differentiation by exchanging ActivinA and XAV939 for 1 µM Chir99021 (a Wnt agonist) and varying concentrations of BMP4 (0-32 ng/ml) and FGF ligands (FGF2 or FGF4).
- Analysis: Assess differentiation outcomes after 3 days using quantitative PCR for markers (Hand1, Gata6, Tbx6, Msgn1, Foxa2, Shh), immunostaining, or single-cell RNA sequencing.
Micropatterned hPSC Systems [10]
- Patterned Culture: Plate human pluripotent stem cells on micropatterned substrates to control colony geometry.
- BMP4 Treatment: Apply BMP4 to induce self-organized patterning into concentric rings of different fates.
- Live Imaging: Monitor signaling dynamics using endogenously tagged GFP::SMAD4 or RFP::SMAD1.
- Endpoint Staining: Fix and stain for fate markers to correlate signaling histories with cell fate decisions.
Genetic Manipulation Approaches
Mutant Analysis [13]
- Zygotic vs. Maternal-Zygotic Mutants: Generate zygotic mutants (deficient in embryonic gdf3) and maternal-zygotic (MZ) mutants (deficient in both maternal and zygotic gdf3) to distinguish maternal and zygotic contributions.
- Rescue Experiments: Express gdf3 and nodal singly or in combination in MZ mutants to test functional requirements.
- Co-immunoprecipitation: Validate heterodimer formation by co-expressing tagged versions of Nodal and Gdf3 and assessing physical interaction.
Single-Cell Analysis of Signaling States
Single-Cell RNA Sequencing [9]
- Differentiation Time Course: Collect cells at multiple time points during EpiSC differentiation.
- Library Preparation and Sequencing: Prepare single-cell libraries using standard platforms (10X Genomics).
- Data Integration: Integrate with published embryonic datasets to map in vitro differentiation to in vivo developmental trajectories.
- Pathway Activity Inference: Calculate pathway activity scores based on expression of known target genes.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Studying Nodal Signaling
| Reagent/Category | Specific Examples | Function/Application |
|---|---|---|
| Signaling Agonists | Recombinant Nodal, ActivinA, BMP4, FGF2/FGF4, Chir99021 | Activate specific signaling pathways in differentiation protocols |
| Signaling Antagonists | XAV939 (Wnt inhibitor), Lefty, Cerberus, SB431542 (Nodal inhibitor) | Inhibit pathway activity to test functional requirements |
| Cell Lines | EpiSCs, mouse ESCs, hPSCs, SBR reporter (T/Bra:mCherry) | Model embryonic development and monitor differentiation in real-time |
| Genetic Tools | gdf3 mutants (zygotic and MZ), CRISPR/Cas9 editing, GFP::SMAD4, RFP::SMAD1 | Manipulate and visualize signaling components |
| Analysis Methods | Single-cell RNA-seq, iterative immunofluorescence, In-Cell Western | Quantify differentiation outcomes and signaling activity |
| Model Systems | Micropatterned hPSCs, 2D gastruloids, 3D aggregate cultures | Study self-organization and patterning in controlled environments |
Nodal signaling represents a paradigm for how morphogens orchestrate complex developmental processes through precise concentration-dependent effects, intricate feedback regulation, and sophisticated integration with other signaling pathways. The recent recognition that Nodal functions predominantly as a heterodimer with Gdf3 resolves long-standing questions about Gdf3 function while revealing new layers of regulation in embryonic patterning. The dynamic, time-integrated nature of signaling responses explains how stable cell fate decisions emerge from transient signaling states.
Future research directions will likely focus on quantifying signaling dynamics at higher spatiotemporal resolution, modeling the complete gene regulatory networks controlled by Nodal, and exploiting this knowledge for directed differentiation of stem cells into specific therapeutic cell types. The continued development of sophisticated embryo models will further bridge the gap between in vitro signaling studies and in vivo embryonic development, providing a more comprehensive understanding of how Nodal establishes the embryonic axis and patterns the mesendoderm during mammalian gastrulation.
Gastrulation is a foundational process in mammalian development, transforming a simple embryonic structure into a complex multi-layered organism. During this critical phase, Fibroblast Growth Factor (FGF) signaling operates in concert with BMP, Nodal, and Wnt pathways to coordinate cell fate specification and morphogenetic movements [3] [16]. In mouse gastrulation, these signaling pathways collectively induce and pattern the primitive streak, serving as the central organizing structure where epiblast cells undergo epithelial-to-mesenchymal transition (EMT) and begin forming the mesoderm and endoderm layers [3] [12].
The FGF pathway emerges as particularly crucial for regulating cell movements during gastrulation. While all these pathways have recognized instructive roles in cell fate specification, growing evidence indicates that BMP, Nodal, and FGF signaling also directly regulate cell movements through mechanisms distinct from those specifying cell fates [16]. This dual functionality enables these pathways to coordinate tissue patterning with the complex morphogenetic events that shape the embryonic body plan.
Molecular Mechanisms of FGF-Regulated EMT
Core FGF Signaling Components and Downstream Effectors
FGF signaling during mouse gastrulation primarily occurs through FGFR1 receptors that activate multiple intracellular pathways. The core molecular machinery involves:
- Ligand-Receptor Interaction: FGF ligands (particularly FGF4 and FGF8) bind to and activate FGFR1 tyrosine kinase receptors in the primitive streak region [17] [16]
- Downstream Pathway Activation: Ligand-bound receptors initiate both the RAS/MAPK and PI3K/AKT signaling cascades [17]
- Transcriptional Regulation: FGF signaling activates expression of the Snail transcriptional repressor (Snai1), which directly represses E-cadherin expression [16]
This molecular cascade directly facilitates EMT by dismantling epithelial characteristics and activating mesenchymal genes essential for cell migration.
E-Cadherin Regulation: Mouse vs. Chicken Models
The mechanism of E-cadherin regulation during FGF-mediated EMT demonstrates significant variation between model organisms:
Table: Comparative Mechanisms of FGF-Regulated EMT
| Aspect | Mouse Model | Chicken Model |
|---|---|---|
| E-cadherin Regulation | Transcriptional repression via Snail [16] | No significant change in expression or localization [17] |
| EMT Requirement | Essential for mesodermal progenitor ingression [16] | Required but through E-cadherin-independent mechanisms [17] |
| Primary FGF Receptors | FGFR1 [16] | FGFR1 (primary), with FGFR2/3 in lateral epiblast [17] |
| Cell Migration Defect | Failed ingression without E-cadherin downregulation [16] | Blocked mesoderm formation despite normal E-cadherin [17] |
This comparative analysis reveals evolutionary divergence in how FGF signaling executes the conserved gastrulation program across species, highlighting the importance of validating findings across multiple experimental models.
FGF Signaling in Mesoderm Migration
Guidance of Mesodermal Progenitors
Following EMT, FGF signaling provides critical guidance cues that direct the migration of mesodermal cells away from the primitive streak. In both mouse and chicken embryos, FGF ligands function as chemotactic signals that spatially organize mesoderm migration [16]:
- FGF8 expressed in the primitive streak acts as a chemorepellant, directing migration of mesodermal cells away from the streak region
- FGF4 expressed in the extending axial mesoderm serves as a chemoattractant for dorsal convergence of lateral mesoderm
- This chemotactic function establishes a migration guidance system that properly positions mesodermal populations for subsequent differentiation
The critical nature of this guidance system is demonstrated in mouse embryos lacking Fgf8, where mesoderm cells gastrulate but fail to migrate away from the primitive streak, resulting in severe developmental defects [17].
FGF Signaling in Germ Cell Migration
Beyond mesoderm formation, FGF signaling also plays specialized roles in guiding primordial germ cells (PGCs) during mouse embryogenesis. Research demonstrates that migrating germ cells express Fgfr1-IIIc and Fgfr2-IIIb receptors and exhibit FGF-mediated MAP kinase pathway activation during their migration [18].
Functional studies reveal distinct roles for different FGF ligands in germ cell behavior:
- FGF2 (ligand for FGFR1-IIIc) directly affects germ cell motility
- FGF7 (ligand for FGFR2-IIIb) primarily regulates germ cell numbers
- Inhibition of FGF signaling causes increased apoptosis and inhibits cell shape changes in migrating germ cells
Genetic evidence from FGFR2-IIIb deficient embryos confirms that FGF signaling through this receptor is essential for maintaining proper germ cell populations, though migration appears unaffected in this specific context [18].
Quantitative Analysis of FGF Signaling Effects
Experimental Inhibition of FGF Signaling
Studies quantifying the effects of FGF signaling inhibition during gastrulation reveal its profound impact on cell migration:
Table: Quantitative Effects of FGFR Inhibition on Cell Migration
| Embryonic Region | DMSO Control (%) | SU5402 Treatment (%) | P-value |
|---|---|---|---|
| Epiblast (Lateral + Preingression) | 59.0 | 78.5 | <0.001 |
| Primitive Streak | 13.4 | 20.6 | <0.001 |
| Mesoderm Layer | 27.6 | 1.0 | <0.001 |
These data demonstrate that pharmacological inhibition of FGFR activity with SU5402 virtually abolishes cell migration into the mesoderm layer, confirming FGF signaling as essential for mesoderm formation in chicken embryos [17]. Similar approaches in mouse models show parallel requirements, though through different mechanistic pathways.
microRNA Regulation by FGF Signaling
FGF signaling additionally regulates gastrulation through post-transcriptional mechanisms involving microRNAs:
Table: FGF-Regulated microRNAs and Their Targets
| microRNA | Regulation Mechanism | Validated mRNA Targets | Functional Impact |
|---|---|---|---|
| let-7b, miR-19b, miR-130b | LIN28B-dependent processing [19] | PDGFRA, TGFBR1 [19] | Cell migration through primitive streak |
| miR-9, miR-107, miR-218 | LIN28B-independent [19] | ACVR1, ACVR2B, TGFBR3 [19] | Mesoderm formation and patterning |
| miR-130b, miR-218 | Direct FGF regulation [19] | PDGFRA [19] | Receptor tyrosine kinase signaling modulation |
This microRNA regulatory network represents a novel mechanism by which FGF signaling fine-tunes gene expression during gastrulation, with LIN28B serving as a critical intermediary in both dependent and independent pathways [19].
Experimental Approaches and Methodologies
Key Research Models and Reagents
The study of FGF signaling in gastrulation employs diverse experimental models, each offering unique advantages:
Essential Research Reagents and Tools
Table: Key Experimental Reagents for FGF Signaling Research
| Reagent/Tool | Application | Function/Mechanism |
|---|---|---|
| SU5402 | FGFR inhibition [19] [17] | Competitive inhibitor of FGFR tyrosine kinase activity at 100 μM concentration |
| LIN28B morpholinos | LIN28B knockdown [19] | Antisense to exon-intron boundary of LIN28B exon 2 to block splicing |
| pBE-LIN28B plasmid | LIN28B overexpression [19] | Chicken β-actin promoter driving LIN28B-FLAG expression |
| GFP expression plasmids | Cell lineage tracing [17] | Electroporation-based labeling to track cell movements |
| U0126 MEK inhibitor | ERK pathway inhibition [20] | Selective blockade of MAPK/ERK signaling downstream of FGFR |
| miR mimics | microRNA functional studies [19] | Gain-of-function analysis of specific FGF-regulated microRNAs |
Detailed Methodological Protocols
Chicken Embryo Electroporation and Migration Assay
This protocol enables precise analysis of cell migration through the primitive streak:
- Embryo Preparation: Incubate fertile chicken eggs until embryos reach Hamburger-Hamilton stage 4 (approximately 24 hours) [19] [17]
- Experimental Treatment: Pretreat embryos for two hours with either:
- 100 μM SU5402 (FGFR inhibitor) dissolved in DMSO
- DMSO alone as vehicle control [17]
- Electroporation: Introduce GFP expression plasmid specifically into epiblast cells using optimized electroporation parameters [17]
- Migration Analysis: Incubate embryos for five hours at 37°C, then fix and analyze GFP-positive cell distributions across:
- Lateral and preingression epiblast
- Primitive streak region
- Mesoderm layers [17]
- Quantification: Count GFP-positive cells in each region and calculate percentage distributions; compare experimental and control conditions using appropriate statistical tests (e.g., Student's t-test) [17]
microRNA Expression Analysis
Comprehensive profiling of FGF-regulated microRNAs involves multiple complementary approaches:
- RNA Isolation: Extract total RNA from primitive streak regions using mirVana miR isolation kit [19]
- microRNA Microarray:
- Label RNA samples with Hy3/Hy5 fluorescent dyes using miRCURY LNA microRNA power labeling kit
- Hybridize to multispecies arrays based on miRBase version 11
- Process slides and analyze data with statistical computing software [19]
- Validation by qRT-PCR:
- Synthesize cDNA using NCode miR first-strand cDNA synthesis kit
- Perform real-time PCR with miR-specific primers
- Normalize using snRNA U6 as internal control [19]
- In Situ Hybridization:
- Fix embryos in 4% paraformaldehyde
- Perform whole-mount ISH using LNA probes labeled with digoxigenin
- Detect specific miR spatial expression patterns [19]
Integrated Signaling Dynamics in Gastrulation
Interpathway Crosstalk and Coordination
FGF signaling does not function in isolation but participates in extensive crosstalk with other major gastrulation pathways:
This integrated signaling network creates a robust regulatory system that ensures proper coordination between cell fate specification and morphogenetic movements. Recent studies using human pluripotent stem cell (hPSC) models confirm that BMP, Nodal, and FGF signaling function in a transcriptional hierarchy during self-organized pattern formation, with each pathway controlling distinct but overlapping aspects of gastrulation [10].
Temporal Dynamics of Signaling Activity
Emerging evidence emphasizes the importance of signaling duration rather than just concentration in determining cell fates. Research in hPSC gastruloids demonstrates that:
- BMP response varies more strongly in duration than in level
- Both level and duration of signaling activity control cell fate choices by changing the time integral of signaling
- Signaling duration and level are interchangeable parameters in this context [10]
This temporal dimension adds complexity to our understanding of how FGF signaling patterns the embryo, suggesting that cells integrate signaling activity over time rather than simply responding to instantaneous concentrations.
FGF signaling serves as a master regulator of gastrulation, coordinating both the EMT process that releases cells from the epiblast and their subsequent migration to proper embryonic positions. Through its interplay with BMP, Nodal, and Wnt pathways, FGF signaling ensures the precise spatial and temporal coordination of cell fate specification and morphogenesis.
The mechanistic insights gained from studying FGF signaling in mouse gastrulation have broader implications for understanding human development and disease. Furthermore, the principles of FGF-mediated cell guidance may inform regenerative medicine approaches aimed at controlling stem cell migration and differentiation in therapeutic contexts.
As research continues to unravel the complexities of FGF signaling, particularly through advanced models like gastruloids and live imaging technologies, our understanding of how this pathway integrates with other signals to shape the embryonic body plan will continue to deepen, offering new insights into both normal development and pathological processes.
The transformation of a simple embryonic structure into a complex, elongated body plan is a cornerstone of vertebrate development. This process, gastrulation, is governed by a complex interplay of morphogen signals that provide positional information to cells. Among these, Bone Morphogenetic Protein (BMP), Nodal, and Fibroblast Growth Factor (FGF) pathways are critical. Recent research underscores that it is not merely the presence of these signals, but their precise spatiotemporal dynamics and relative ratios that drive the distinct cellular behaviors of convergence and extension (C&E) gastrulation movements in different tissues [21] [22]. This whitepaper synthesizes current findings on how the dynamics of BMP and Nodal signaling orchestrate tissue-specific morphogenesis, with a specific focus on insights from mouse and zebrafish model systems relevant to the broader thesis of signaling pathways in mouse gastrulation.
Core Findings: Temporal Dynamics of Morphogen Ratios Drive Tissue-Specific Morphogenesis
A pivotal concept emerging from recent studies is that the morphogenetic outcome of a cell is not determined by a single morphogen in isolation, but by the dynamic balance between multiple pathways over time.
The BMP/Nodal Ratio as a Master Regulator
Using zebrafish explant models, researchers have demonstrated that the ratio of BMP to Nodal signaling activity during a critical developmental window dictates whether morphogenesis proceeds via neuroectoderm-driven or mesoderm-driven C&E [22].
- High BMP/Nodal Ratio: Promotes neuroectoderm (NE)-driven convergent extension. In this regime, C&E movements are primarily executed by cells of the neuroectoderm [21] [22].
- Low BMP/Nodal Ratio: Promotes mesoderm-driven convergent extension. Here, the underlying mesodermal tissue is the primary driver of the narrowing and elongation of the embryonic axis [22].
This mechanistic insight reveals that the same core signaling pathways can activate distinct morphogenetic programs within different tissues based on their relative levels and timing.
Critical Windows of Signaling Activity
The ability of the BMP/Nodal ratio to specify the mode of morphogenesis is confined to a specific temporal window during gastrulation. Precise optogenetic manipulation of signaling onset has been used to identify these critical windows, demonstrating that the competence of tissues to respond to morphogen cues is tightly regulated in time [22].
Table 1: Summary of Morphogen Signaling Effects in Gastrulation
| Signaling Pathway | Primary Role in Gastrulation | Effect on Convergent Extension (C&E) | Key Interactions |
|---|---|---|---|
| Nodal | Mesoderm specification; promotes gastrulation movements [22]. | Promotes C&E cell behaviors cell-autonomously in both mesoderm and neuroectoderm [22]. | Ratio with BMP dictates tissue-specific C&E mode; target gene chordin is a BMP antagonist [22]. |
| BMP | Specifies ventral embryonic cell types [22]. | High levels generally inhibit C&E; a high BMP/Nodal ratio specifically promotes NE-driven C&E [21] [22]. | Antagonized by Nodal-induced Chordin; ratio with Nodal is a key determinant of morphogenetic outcome [22]. |
| Wnt/β-catenin | Axis specification; posterior patterning [23]. | Essential for symmetry breaking and axis elongation; posterior pole of Wnt activity defines the A-P axis [23]. | Interacts with Nodal and BMP pathways; its polarization is preceded by Nodal and BMP heterogeneity [23]. |
Experimental Models and Methodologies
Investigating spatiotemporal morphogen dynamics requires sophisticated model systems and precise experimental techniques.
Key Model Systems
- Zebrafish Explants ("Pescoids"): Relatively naïve clusters of cells from the animal pole of zebrafish embryos can be induced to undergo C&E with exogenous Nodal pathway activation. These explants allow for the physical and chemical isolation of tissues, enabling the study of NE morphogenesis uncoupled from the mesoderm [22].
- Mouse Gastruloids: These 3D aggregates of mouse embryonic stem cells (mESCs) break symmetry and self-organize into structures with an anterior-posterior (A-P) axis, mimicking aspects of mammalian gastrulation. They provide a powerful platform for studying self-organization without extrinsic embryonic cues [23].
Advanced Techniques for Tracing Morphogen Signals
A breakthrough in understanding the temporal evolution of signaling patterns has come from the development of synthetic "signal-recording" gene circuits [23].
Detailed Protocol: Signal-Recording Gene Circuit in mESCs [23]
Circuit Design: An AND-gate gene circuit is engineered into mouse ESCs. This circuit consists of:
- A "sentinel enhancer" responsive to a specific morphogen pathway (e.g., TCF/LEF for Wnt signaling).
- A destabilized, doxycycline (dox)-dependent transcription factor (rtTA) expressed downstream of the sentinel enhancer.
- An rtTA-dependent promoter (PTetON) driving expression of a destabilized Cre recombinase.
- A reporter gene (e.g., dsRed) that is permanently switched to another fluorescent protein (e.g., GFP) upon Cre-mediated recombination.
Mechanism of Action: Only when BOTH the morphogen pathway is active AND doxycycline is present in the media will rtTA activate the PTetON promoter. This leads to Cre expression and a permanent, heritable switch in the fluorescent reporter from dsRed to GFP. The progeny of the labeled cell will inherit the GFP signal.
Temporal Control: By pulsing doxycycline for as little as 1.5–3 hours, researchers can permanently "record" which cells were experiencing active morphogen signaling during that specific time window.
Application: This technique was used to trace the origins of the Wnt-active posterior pole in mouse gastruloids. It revealed that patchy domains of Wnt-high cells, which emerge from earlier Nodal and BMP heterogeneity, rearrange via cell sorting to form a single, coherent polarized domain [23].
The following diagram illustrates the workflow and logic of this signal-recording technique:
Single-Embryo, Single-Cell Resolution
Advances in single-cell RNA sequencing (scRNA-seq) have enabled the creation of high-resolution temporal models of mouse gastrulation. One study profiled 153 individually sampled embryos, allowing inference of differentiation flows and lineage specification dynamics. This approach revealed that gastrulation is dominated by progenitor states that undergo continuous multi-furcation rather than a simple series of binary fate decisions, highlighting the complex interplay of transcription factors in regulating these processes [24].
The Scientist's Toolkit: Essential Research Reagents
The following table details key reagents and tools used in the cited experiments to study morphogen dynamics.
Table 2: Key Research Reagents for Investigating Morphogen Dynamics
| Research Reagent | Function & Application in Research |
|---|---|
| Zebrafish Animal Pole Explants | A model system of naïve embryonic cells used to study tissue-specific morphogenesis uncoupled from endogenous signals; induced with Nodal to study NE-vs-mesoderm C&E [22]. |
| Constitutively Active Nodal Receptor (CA-acvr1b*) | A tool for robust, early activation of the Nodal signaling pathway; in explants, induces mesoderm-driven convergent extension [22]. |
| Synthetic Signal-Recording Gene Circuits | Engineered genetic constructs that permanently record a cell's historical exposure to a specific morphogen signal (e.g., Wnt, Nodal) during a user-defined time window (controlled by doxycycline) [23]. |
| Optogenetic Manipulation Systems | Allows precise temporal control over the onset of signaling pathways (e.g., Nodal) to identify critical developmental windows for morphogen function [22]. |
| lhx1a:EGFP Transgenic Zebrafish | A transgenic line where the axial and lateral/intermediate mesoderm are labeled with EGFP; enables visualization of mesoderm formation and positioning in explants and intact embryos [22]. |
| PTCF/LEF-iRFP-PEST Biosensor | A live-cell biosensor reporting real-time Wnt/β-catenin pathway activity through the expression of a destabilized infrared fluorescent protein [23]. |
Integrated Signaling Pathway and Experimental Workflow
The relationship between key morphogens and a core experimental method is summarized in the following pathway diagram, which integrates the roles of Nodal, BMP, and Wnt with the signal-recording workflow.
The formation of the vertebrate body plan is an exquisitely coordinated process driven by the spatiotemporal dynamics of morphogen distribution. The evidence is clear that the temporal dynamics of the BMP/Nodal ratio act as a critical switch directing distinct, tissue-specific morphogenetic programs during gastrulation. The emergence of sophisticated tools—including stem cell-derived gastruloid models, synthetic biology approaches for recording signaling history, and single-cell genomics—is providing an unprecedented view into these processes. Understanding these dynamics is not only fundamental to developmental biology but also provides critical insights for regenerative medicine and drug development, where manipulating cell fate and tissue organization is paramount. Future research will continue to unravel how these signaling pathways are integrated in time and space to build a complex organism from a simple cluster of cells.
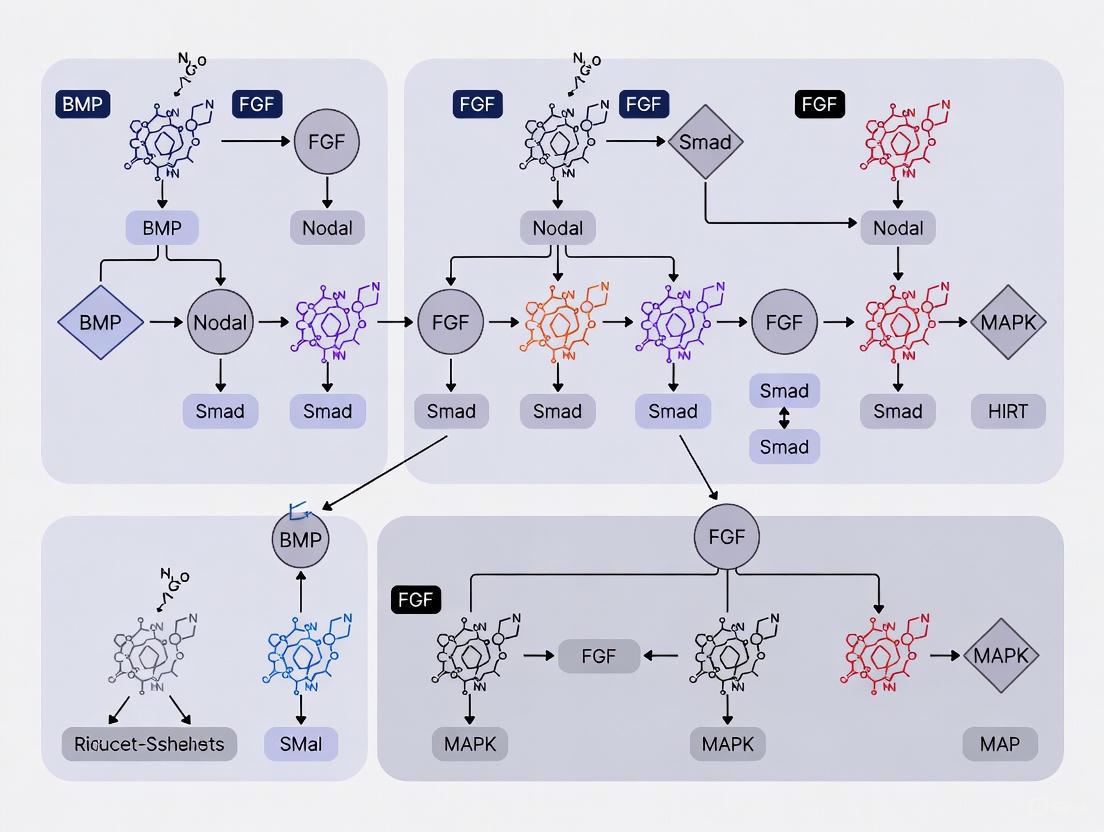
The formation of the mammalian body plan during gastrulation is directed by a complex, self-organizing signaling network. Core pathways—BMP, Wnt, Nodal, and FGF—do not operate in isolation but engage in extensive crosstalk, including reciprocal activation, inhibition, and synergistic interactions that ensure precise spatial and temporal control of cell fate specification. This whitepaper synthesizes current research to present an integrated view of this network. It details the regulatory logic governing pathway interactions, provides quantitative data on signaling inputs and fate outputs, and outlines key experimental methodologies for its study. Framed within the context of mouse gastrulation, this resource is designed to equip researchers and drug development professionals with the mechanistic insights and tools necessary to interrogate and manipulate this fundamental biological system.
Gastrulation is a transformative period in mouse embryonic development whereby pluripotent cells of the epiblast exit the naive state and undergo coordinated movements and differentiation to form the three definitive germ layers: the ectoderm, mesoderm, and endoderm. This process is orchestrated by embryonic and extraembryonic signaling centers that release a tightly controlled combination of secreted ligands, establishing dynamic signaling environments [25]. The convergence of Bone Morphogenetic Protein (BMP), Wnt, Nodal, and Fibroblast Growth Factor (FGF) signaling is the principal driver of gastrulation. Disruption of any single pathway results in catastrophic failure of gastrulation and embryonic lethality, underscoring their non-redundant and critical functions [25].
The primitive streak, the site of gastrulation, is induced and patterned by these pathways. Cell fate decisions are not dictated by a single signal but are instead a product of the specific combination, level, and duration of signals a cell perceives, which varies according to its position within the embryo and the developmental time window [25]. Historically, genetic experiments in mouse embryos have identified the essential components. However, the intricate dynamics of how these pathways interact as an integrated network have been difficult to dissect in vivo due to the complexity of the embryonic architecture and technical limitations. The recent advent of sophisticated in vitro stem cell models, such as gastruloids and patterned epiblast stem cell (EpiSC) cultures, has provided a scalable, tractable, and deconstructed system to probe these complex interactions with high resolution [12] [9]. These models have begun to reveal a core regulatory logic, characterized by community effects, positive autoregulation, and pathway antagonism, which ensures the robust and reproducible patterning of the early embryo.
The Core Signaling Network: Components and Interactions
The signaling network operates through a precise spatial organization. The proximal-to-distal axis of the embryo is patterned by a gradient of BMP signaling, highest proximally due to ligand expression from the extraembryonic ectoderm (ExE). This is complemented by Wnt and Nodal gradients, which are strongest posteriorly and are antagonized by inhibitors from the anterior visceral endoderm (AVE) [9]. FGF signaling is most active within and surrounding the primitive streak and nascent mesoderm [9]. The following diagram illustrates the key interactions and regulatory relationships between these core pathways.
Figure 1: Core Signaling Network in Mouse Gastrulation. Pathways activate (solid arrows) or inhibit (dashed arrows) biological processes and each other. FGF exhibits positive autoregulation and repression of BMP, while Wnt and Nodal can cross-activate the FGF pathway.
Key Regulatory Logic and Antagonisms
The network is defined by several core regulatory motifs that ensure robust patterning.
BMP-FGF Antagonism: A fundamental axis of interaction is the opposing relationship between BMP and FGF. BMP signaling promotes proximal mesoderm fates (e.g., extraembryonic mesoderm, blood precursors). In contrast, FGF signaling is essential for the specification and migration of distal mesoderm fates (e.g., paraxial and axial mesoderm) [9]. Single-cell RNA sequencing analyses of differentiating EpiSCs reveal that FGF stimulation not only boosts the expression of endogenous Fgf genes, creating a positive feedback loop, but also actively represses the expression of Bmp ligand genes [9]. This dual mechanism establishes a coherent, self-reinforcing signaling domain for distal fates while simultaneously suppressing the competing BMP-dependent proximal fate program.
Cross-Activation Constrains Cell States: The network's interconnectedness also serves to constrain cell fate potential. Research in mouse Embryonic Stem Cells (mESCs) has shown that BMP signaling can induce a totipotent state, reminiscent of the pre-implantation embryo. However, this induction is naturally limited because BMP activation leads to the cross-activation of FGF, Nodal, and Wnt pathways. The activity of these cross-activated pathways subsequently constrains the expansion of the totipotent cell population. Only by rationally inhibiting all three cross-activated pathways can the efficiency of BMP-mediated totipotency induction be significantly enhanced [26]. This demonstrates how cross-pathway interactions maintain heterogeneity and prevent aberrant state transitions.
Quantitative Signaling-Fate Relationships
Cell fate is determined by the specific concentration and combination of signaling inputs. The tables below summarize quantitative data from in vitro EpiSC differentiation studies, mapping signaling inputs to the expression of key marker genes and the resulting mesodermal subtypes.
Table 1: Concentration-Dependent Effects of BMP4 on Mesoderm Marker Expression after 3 Days of EpiSC Differentiation (based on qPCR data) [9]
| BMP4 Concentration (ng/ml) | Marker Gene Expression (Peak) | Representative Embryonic Region |
|---|---|---|
| 0 - 2 | Foxa2, Shh (Distal markers) | Anterior Primitive Streak / Endoderm |
| ~4 | T/Bra, Tbx6, Msgn1 | Mid/Distal Mesoderm (Paraxial) |
| 16 - 32 | Hand1, Gata6 (Proximal markers) | Posterior Proximal Mesoderm (Extraembryonic, Heart) |
Table 2: FGF Signaling Dependence of Major Mesodermal Lineages [9]
| Mesodermal Lineage | Dependence on FGF Signaling | Key Markers | Notes |
|---|---|---|---|
| Extraembryonic Mesoderm | FGF-independent | Hand1 | Forms in FGF receptor mutants. |
| Cardiac & Head Mesoderm | Partially FGF-dependent | Mesp1 | May require low-level FGF. |
| Paraxial & Axial Mesoderm | FGF-dependent | Tbx6, Msgn1 | Deficient in FGF pathway mutants; requires FGF for robust specification. |
Experimental Protocols for Network Analysis
The following section outlines key methodologies used to dissect the integrated signaling network, leveraging the power and scalability of in vitro stem cell models.
2D Epiblast Stem Cell (EpiSC) Differentiation and Signaling Titration
This protocol allows for the systematic analysis of how specific signaling pathway concentrations influence mesoderm differentiation in a homogeneous cell population [9].
Cell Culture and Maintenance: Maintain EpiSCs (e.g., SBR reporter line for T/Bra expression) in FAX medium: N2B27 base medium supplemented with ActivinA (to maintain pluripotency), FGF2 (e.g., 12 ng/ml), and the Wnt signaling inhibitor XAV939 (to suppress heterogeneity). Cells should be kept in a homogeneous, pluripotent state (NANOG-positive, T/BRA-negative).
Induction of Mesoderm Differentiation: To trigger differentiation, passage cells and change to a medium containing the Wnt agonist CHIR99021 (Chi, 1 µM) and varying concentrations of the signaling factors under investigation (e.g., BMP4 and/or FGF2/FGF4). The standard BMP4 concentration for generating a mixed population is 8 ng/ml.
Signaling Titration: To determine concentration-dependent effects, set up parallel differentiation cultures with a titration series of the ligand of interest (e.g., BMP4 from 0 to 64 ng/ml, or FGF2 from 0 to 100 ng/ml).
Analysis (After 72 hours of differentiation):
- Quantitative PCR (qPCR): Harvest cells for RNA extraction and analyze expression of a panel of region-specific markers (e.g., Hand1, Gata6 for proximal; Tbx6, Msgn1 for distal; Foxa2 for anterior).
- In-Cell Western Assay: Fix cells in-plate and perform immunodetection for proteins like HAND1 and GATA6. This allows for quantitative, high-throughput protein-level analysis across the titration series.
- Flow Cytometry: If using reporter lines like T/Bra:mCherry, analyze the percentage of positive cells and fluorescence intensity to quantify differentiation efficiency.
Single-Cell RNA Sequencing Analysis
To deconstruct the heterogeneity of the differentiated populations and precisely map FGF-dependent and -independent lineages, single-cell RNA sequencing (scRNA-seq) is employed [9].
Sample Preparation: Differentiate EpiSCs under control and FGF-inhibited conditions (e.g., using an FGFR inhibitor). After 3 days, create a single-cell suspension.
Library Preparation and Sequencing: Use a platform like the 10x Genomics Chromium system to barcode and prepare libraries from thousands of individual cells. Sequence the libraries to obtain transcriptome data for each cell.
Bioinformatic Analysis:
- Preprocessing and Clustering: Process raw data (quality control, normalization) and use dimensionality reduction (PCA, UMAP) and clustering algorithms (e.g., Seurat, Scanpy) to identify distinct cell populations.
- Cell Type Annotation: Identify the identity of each cluster by comparing the expression of known marker genes to reference datasets from the gastrulating mouse embryo.
- Differential Expression and Pathway Analysis: Compare clusters from different conditions to identify FGF-dependent gene programs. Analyze the expression of ligand and receptor genes to infer pathway activity (e.g., confirmed FGF-mediated repression of Bmp4).
Computational Consensome Analysis
The Signaling Pathways Project (SPP) knowledgebase enables the prediction of downstream genomic targets of signaling pathway nodes through a meta-analysis technique called consensome analysis [27].
Data Biocuration: SPP biocurates over 10,000 public transcriptomic (e.g., RNA-seq) and cistromic (ChIP-seq) datasets. Each dataset is classified according to the signaling pathway node (e.g., receptor, kinase, transcription factor) that was genetically or pharmacologically perturbed.
Consensome Generation: For a given node family (e.g., BMP receptors, SMAD transcription factors), the pipeline aggregates all relevant transcriptomic datasets. It then ranks genes based on a consensus measure of their significant differential expression across these independent studies.
Hypothesis Generation: Researchers can query the SPP webbrowser (Ominer) to access these pre-computed consensomes. A high rank for a specific gene indicates strong consensus evidence for its regulation by that signaling node family, providing a data-driven starting point for experimental validation.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Investigating Signaling Pathways in Gastrulation Models
| Reagent / Tool | Function / Target | Example Use Case | Key Findings Enabled |
|---|---|---|---|
| Recombinant BMP4 | BMP pathway agonist; binds BMP receptors. | Titration in EpiSC differentiation. | Established concentration-dependent fate specification (proximal vs. distal) [9]. |
| CHIR99021 | Small molecule agonist of Wnt signaling (GSK3 inhibitor). | Induction of primitive streak-like formation in EpiSCs and gastruloids. | Key for initiating the gastrulation program in vitro [9]. |
| FGF2 / FGF4 | Recombinant ligands; activate FGF receptor tyrosine kinases. | Titration to assess FGF-dependent lineages; maintenance of EpiSC pluripotency. | Identified opposing functions to BMP and requirement for distal mesoderm [9]. |
| XAV939 | Small molecule inhibitor of Wnt signaling (Tankyrase inhibitor). | Used in EpiSC maintenance medium to suppress spontaneous differentiation. | Allows for a homogeneous, pluripotent starting population [9]. |
| T/Bra:mCherry Reporter Cell Line | Fluorescent reporter for pan-mesodermal marker T/Brachyury. | Flow cytometry or live imaging to quantify mesoderm differentiation. | Enabled quantitative tracking of mesoderm induction in response to signaling perturbations [9]. |
| FGFR Inhibitors (e.g., PD173074) | Small molecule inhibitors of FGF receptor tyrosine kinases. | To block FGF signaling during EpiSC differentiation. | Mapped FGF-dependent and -independent mesodermal lineages via scRNA-seq [9]. |
Integrated Workflow for Signaling Network Dissection
The following diagram outlines a logical workflow that integrates the protocols and reagents described above to systematically dissect cross-pathway interactions.
Figure 2: Workflow for Dissecting Signaling Networks. A logical pipeline from establishing a controlled starting population to generating a refined network model through iterative perturbation and multi-modal analysis.
The study of signaling during mouse gastrulation has evolved from a focus on individual pathway functions to an appreciation of a deeply interconnected network. The core logic of this network, defined by key antagonisms (BMP vs. FGF) and cross-activations (BMP→FGF/Nodal/Wnt), provides a robust system for patterning the embryo and controlling cell state transitions. The quantitative relationships between signal concentration and fate output, now measurable with high precision in stem cell models, offer a predictive framework for directing cell differentiation for therapeutic purposes.
Future research will be driven by the integration of multi-omics data (transcriptomics, proteomics, phosphoproteomics) and the application of computational modeling [12] [28]. Projects like the Signaling Pathways Project demonstrate the power of aggregating public 'omics data to predict node-target relationships. Meanwhile, the application of similar meta-analysis and machine learning approaches to data generated from controlled in vitro models like gastruloids will be crucial for building dynamic, quantitative models of the network. These models will not only enhance our fundamental understanding of embryogenesis but will also accelerate drug discovery by revealing how complex signaling networks are rewired in developmental diseases and cancer.
Research Tools: From Embryo Models to Single-Cell Omics
Mouse Embryo Staging and Tissue Dissection Strategies
The house mouse (Mus musculus) serves as a premier model system for mammalian developmental biology due to its genetic tractability and close evolutionary affinity to humans [29]. Mouse gestation encompasses a remarkable transformation, from a single-cell zygote to a free-living pup composed of more than 500 million cells in just three weeks [29]. Within this compact timeline, the process of gastrulation represents a critical period during which the three definitive germ layers—ectoderm, mesoderm, and endoderm—are established. This in-depth technical guide provides researchers with current methodologies for mouse embryo staging and tissue dissection, with particular emphasis on their application within signaling pathway research, specifically focusing on BMP, Nodal, and FGF pathways during gastrulation.
Mouse Embryo Staging: Principles and Practices
Staging Based on Morphological Criteria
Precise embryonic staging is fundamental for reproducible research, as gestational age alone is an insufficient indicator of developmental progression. Stochastic differences in fertilization timing, genetic factors, and litter size contribute to significant variation among embryos of identical gestational age [29]. Therefore, morphological criteria provide the most reliable assessment of developmental age.
Table 1: Key Staging Parameters for Early Post-implantation Mouse Embryos
| Developmental Stage | Somite Pairs | Crown-Rump Length | Major Developmental Events |
|---|---|---|---|
| Early Gastrulation | 0-5 | Data not specified | Onset of gastrulation, primitive streak formation [1] |
| Mid Gastrulation | 6-12 | Data not specified | Axis patterning, neural plate formation |
| Late Gastrulation | 13-20 | Data not specified | Somitogenesis progression, organ progenitor specification |
| Early Organogenesis | 21-30 | Data not specified | Initiation of organogenesis, limb bud formation |
For studies focusing on gastrulation, which occurs approximately between embryonic day (E) 6.25 and E9.5, somite counting provides a precise morphological staging method [1] [30] [31]. The development of 67 post-implantation mouse embryos cultured in vitro has been demonstrated to progress at a pace very similar to that of embryos developed in vivo, validating the use of these staging methods across experimental conditions [30] [31].
Advanced Imaging for Developmental Analysis
Recent technological advances have revolutionized our ability to monitor embryonic development. Time-lapse optical coherence microscopy (OCM) has emerged as a powerful label-free technique for monitoring embryo development with micron-level resolution, enabling visualization of cellular and subcellular features without compromising viability [32]. This method allows continuous monitoring of embryos from the one-cell stage to fully hatched blastocysts every 10 minutes over approximately six days within incubator conditions [32].
For transcriptional profiling across developmental time, single-cell combinatorial indexing (sci-RNA-seq3) has enabled the generation of comprehensive atlases. One recent landmark study profiled 12.4 million nuclei from 83 embryos precisely staged at 2- to 6-hour intervals spanning late gastrulation (E8) to birth [29]. This approach provides unprecedented resolution for capturing dynamic changes in cell states during critical developmental windows, including gastrulation.
Signaling Pathways in Mouse Gastrulation
Gastrulation represents a pivotal period when signaling pathways interact to coordinate cell fate decisions and morphogenetic events. The establishment of the anterior-posterior (AP) axis precedes and sets the stage for gastrulation, involving a complex interplay of signaling centers.
Axis Patterning and Pathway Integration
The transformation of a radially symmetrical epiblast into an embryo with distinct AP orientation involves sophisticated signaling cross-talk:
Nodal Signaling: Creates a proximal-distal gradient through conversion of pro-NODAL by FURIN and PACE4 convertases in the extra-embryonic ectoderm (ExE) [1]. Low Nodal levels at the distal pole specify distal visceral endoderm (DVE) cells, which subsequently migrate to become anterior visceral endoderm (AVE) [1].
BMP Signaling: Secreted from the ExE, BMP4 inhibits DVE formation, restricting it to the distal pole [1]. Recent research using embryonic stem cells (ESCs) has identified BMP signaling as an inducer of the totipotent state, though this role is constrained by cross-activation of FGF, NODAL, and WNT pathways [26] [15].
Wnt Signaling: The AVE expresses the Wnt inhibitor DKK1, which restricts WNT3a signaling domains originating from the posterior epiblast and visceral endoderm [1]. This restriction is critical for proper positioning of the primitive streak at the posterior pole.
The following diagram illustrates the intricate signaling network that patterns the anterior-posterior axis during mouse gastrulation:
Diagram Title: Signaling Network in Mouse Gastrulation
Pathway Cross-Activation in Cell Fate Determination
Research using mouse embryonic stem cells (ESCs) has revealed crucial constraints on signaling pathways. BMP signaling induces a totipotent state but simultaneously cross-activates FGF, NODAL, and WNT pathways, which in turn constrain this induction [26] [15]. This cross-activation creates a balancing mechanism that regulates stem cell heterogeneity and fate decisions—principles that likely extend to in vivo development during gastrulation.
Experimental inhibition of these cross-activated pathways enhances the proportion of totipotent cells in culture, providing a strategy for manipulating cell states [26] [15]. Single-cell mRNA sequencing has confirmed that induction of the totipotent state through BMP signaling manipulation is accompanied by suppression of primed and primitive endoderm states [15].
Tissue Dissection Strategies for Gene Expression Analysis
Non-Enzymatic Microsurgical Dissection Technique
For gene expression profiling studies, traditional enzymatic separation methods can alter RNA profiles and introduce cross-contamination. A specialized non-enzymatic microsurgical dissection technique has been developed to address these challenges [33].
Key Steps in the Protocol:
Transcriptional Arrest: Embryonic tissues are immediately placed in RNAlater solution following dissection to block transcriptional activity and protect RNA from degradation [33].
Controlled Rehydration: While RNAlater causes severe dehydration that impedes dissection, established rehydration conditions facilitate tissue separation without compromising RNA integrity [33].
Microsurgical Separation: Using fine instruments, tissues are separated through mechanical dissection alone, without enzymatic treatment. The rehydration step shortens total dissection time to approximately 10 minutes [33].
Quality Validation: Extracted RNA quality is assessed, with excellent results suitable for transcriptional profiling. Tissue-specific gene expression analysis by microarray and RT-PCR confirms minimal cross-contamination [33].
This method has been successfully applied to various embryonic organs, including the challenging rudimentary mammary glands, which are particularly difficult to separate from contiguous ectoderm and strongly adhering mesenchyme [33].
Applications in Signaling Pathway Research
This dissection technique enables precise isolation of specific embryonic regions for analyzing localized signaling pathway activity during gastrulation. Researchers can isolate regions corresponding to signaling centers (e.g., primitive streak, AVE) to profile expression patterns of BMP, Nodal, and FGF pathway components with minimal cross-contamination.
The Scientist's Toolkit: Essential Research Reagents
Table 2: Key Research Reagents for Mouse Embryo Studies
| Reagent/Method | Function/Application | Technical Notes |
|---|---|---|
| RNAlater | Preserves RNA integrity during tissue dissection | Enables non-enzymatic tissue separation; rehydration conditions critical for visibility [33] |
| Time-lapse OCM | Label-free imaging of embryo development | Provides micron-level resolution; suitable for long-term culture monitoring [32] |
| sci-RNA-seq3 | Single-nucleus transcriptional profiling | Enables comprehensive atlas generation; applicable to frozen embryos [29] |
| H2B-mCherry mRNA | Nuclear labeling for live imaging | Electroporation efficiency: ~75% in mouse embryos [34] |
| Light-sheet microscopy | Live imaging with minimal phototoxicity | Dual illumination/detection captures dual views; suitable for long-term imaging [34] |
Mouse embryo staging and dissection methodologies have evolved significantly, enabling unprecedented resolution in studying signaling pathways during gastrulation. The integration of precise morphological staging with advanced imaging techniques and careful dissection approaches provides a powerful framework for investigating the intricate signaling networks involving BMP, Nodal, and FGF pathways. These technical advances continue to deepen our understanding of the fundamental principles governing mammalian development, with implications for regenerative medicine, developmental biology, and therapeutic discovery.
Gastruloids are three-dimensional embryonic organoids formed from aggregates of pluripotent stem cells (PSCs) that recapitulate key aspects of early mammalian embryogenesis in vitro. These self-organizing structures mimic the spatial and temporal events of gastrulation—the fundamental process during which the embryo transforms from a simple ball of cells into a complex, multi-layered structure with established body axes [35] [36]. Unlike mammalian embryos, which develop within the uterus and present significant technical and ethical challenges for research, gastruloids offer unprecedented experimental versatility, scalability, and accessibility for studying developmental dynamics [37]. Their ability to model the intricate signaling crosstalk that coordinates germ layer patterning and cell fate specification makes them particularly powerful for investigating the roles of BMP, Nodal, FGF, and Wnt pathways during the critical developmental window of gastrulation [12] [3]. This technical guide explores the application of gastruloids for dissecting signaling dynamics within the context of a broader thesis on mouse gastrulation.
Signaling Pathways in Gastruloid Self-Organization
The self-organization of gastruloids is orchestrated by a tightly regulated interplay of conserved signaling pathways. Our understanding of this process, historically derived from genetic studies in mouse embryos, has been significantly enhanced by the experimental tractability of gastruloid models [12] [3].
- BMP Signaling: The BMP pathway acts as a key initiator of patterning. In gastruloids, externally applied BMP4 can induce the radial organization of germ layers, mirroring its role in vivo. A critical finding from gastruloid studies is that BMP4 directly induces the expression of its own inhibitor, NOGGIN. This establishes a reaction-diffusion system wherein the diffusion of NOGGIN away from the source creates a gradient that helps define sharp boundaries between different fate domains [38].
- Wnt/β-catenin and Nodal Signaling: Wnt and Nodal are pivotal for axis formation and mesendoderm specification. In standard mouse gastruloid protocols, the transient activation of Wnt signaling (e.g., via CHIR99021) is sufficient to break symmetry and initiate axial elongation, recapitulating the role of the primitive streak [39]. These pathways engage in complex crosstalk, with Nodal signaling reinforcing the Wnt-induced patterning network [12].
- FGF Signaling: The FGF pathway is involved in the epithelial-to-mesenchymal transition (EMT) and subsequent mesodermal patterning. Gastruloid studies have shown that FGF signaling, often in concert with Wnt and Nodal, is required for the proper specification and migration of nascent mesodermal populations [12].
The following diagram illustrates the core signaling network and its functional outcomes in a developing gastruloid:
Quantitative Signaling Dynamics and Morphogen Responses
Gastruloids enable the quantitative measurement of signaling activity and cell fate decisions across space and time. The table below summarizes key quantitative findings from foundational gastruloid studies, highlighting the precision of this model.
Table 1: Quantitative Signaling and Fate Readouts in Gastruloid Models
| Parameter Measured | Experimental Finding | Significance / Implication | Source |
|---|---|---|---|
| BMP4 sensitivity (Kmp) | Edge cells: < 0.5 ng/ml; Center cells: Loss of sensitivity >50 µm from edge | Density-dependent receptor localization creates a pre-pattern for fate specification. | [38] |
| Early hematopoietic progenitor emergence | c-Kit+/CD34+ population upregulated from 120 h; c-Kit+/CD34+/CD41+ population appears after 144 h. | Recapitulates the temporal sequence of embryonic blood development with high fidelity. | [37] |
| Erythroid-like cell emergence | Ter119+ cells appear around 120 h, maintained until 168 h. | Models the onset of primitive erythropoiesis in vitro. | [37] |
| Teratogenicity assessment | Significant reduction in elongation or size; disruption of SOX2/BRA/SOX17 expression patterns. | Provides a quantitative, medium-throughput platform for developmental toxicity screening. | [39] |
Experimental Workflows: From Pluripotency to Patterned Gastruloids
A major strength of the gastruloid model is the highly standardized protocol for generating reproducible, patterned structures from pluripotent stem cells. The workflow below details the key steps for generating mouse gastruloids, which can be adapted for specific signaling studies.
Standard Mouse Gastruloid Protocol
- Cell Culture Maintenance: Mouse embryonic stem cells (mESCs) are maintained in ES medium supplemented with LIF on gelatin-coated tissue culture plastic. Cells should be passaged every two days to ensure a pluripotent, undifferentiated state [39].
- Pre-treatment (Optional): One day prior to aggregation, the culture medium can be changed to 2i/LIF medium to synchronize cells in a ground state of pluripotency [39].
- Aggregation and Initiation:
- A single-cell suspension is prepared, and cells are counted.
- Precisely 300 mESCs are aliquoted in 40 µl of N2B27 medium into each well of a U-bottomed, low-adhesion 96-well plate.
- The plate is centrifuged briefly to encourage aggregate formation at the bottom of each well [39].
- Patterning and Differentiation:
- The aggregates are cultured for 48 hours to form compact, spherical structures.
- At the 48-hour time point, a pulse of Wnt activation (e.g., CHIR99021, a GSK3β inhibitor) is applied for ~24 hours to break symmetry and induce axial organization [39].
- The gastruloids continue to develop in base N2B27 medium or can be steered toward specific fates. For example, to promote cardiovascular and hematopoietic development, VEGF, bFGF, and ascorbic acid can be added from day 3 or 4 onwards [37].
- Analysis and Readout: Gastruloids develop over 120-168 hours, during which they can be analyzed for morphological changes, gene expression (e.g., RNA-seq, qPCR), protein localization (immunofluorescence), and cell composition (flow cytometry) [37] [39].
The following diagram visualizes this standard workflow and its key branching options for different experimental goals:
The Scientist's Toolkit: Essential Reagents and Solutions
Successful gastruloid culture and analysis depend on a suite of well-defined reagents and tools. The following table catalogs essential components for setting up and interrogating signaling in gastruloids.
Table 2: Essential Research Reagent Solutions for Gastruloid Signaling Studies
| Reagent / Tool | Function / Application | Example Use in Gastruloids | |
|---|---|---|---|
| N2B27 Medium | A chemically defined, serum-free basal medium. | Supports unbiased differentiation and is the foundation for all gastruloid culture protocols. | [39] |
| CHIR99021 | A small-molecule inhibitor of GSK3β that activates Wnt/β-catenin signaling. | Used in a pulse (e.g., 48-72h) to break symmetry and induce primitive streak-like fate. | [39] |
| Recombinant BMP4 | A recombinant morphogen protein of the TGF-β superfamily. | Used in micropatterned hESC models to radially pattern germ layers; key for studying BMP-Nodal crosstalk. | [38] |
| VEGF & bFGF | Recombinant growth factors for vascular and fibroblast growth factor signaling. | Added to culture medium to steer gastruloids toward cardiovascular and hematopoietic fates. | [37] |
| Sox1-GFP::Brachyury-mCherry Reporter Line | A dual-reporter mESC line. | Allows real-time live imaging of neuroectoderm (Sox1) and mesoderm (Brachyury) specification in situ. | [37] [39] |
| Fluorescent Reporters (e.g., SOX2, BRA, SOX17) | Transgenic cell lines with fluorescent tags under germ-layer-specific promoters. | Enables quantitative analysis of axial patterning and germ layer proportion upon teratogen or signaling perturbation. | [39] |
Gastruloids have firmly established themselves as a powerful and disruptive platform for studying signaling dynamics during mammalian gastrulation. Their ability to model the self-organized interplay of BMP, Wnt, Nodal, and FGF pathways with spatiotemporal fidelity provides an unparalleled in vitro window into early embryonic events. The quantitative, scalable, and ethically advantageous nature of gastruloids positions them to dramatically accelerate fundamental research in developmental biology and to transform applied fields such as teratogenicity testing and disease modeling. As protocols for generating ever more complex gastruloids continue to evolve, integrating these models with computational approaches will be key to achieving a truly predictive understanding of how signaling networks orchestrate the emergence of form and function in the mammalian embryo.
Spatial Transcriptomics and Epigenomic Profiling of Signaling Components
Spatial transcriptomic technologies represent a revolutionary advance in molecular biology, enabling the comprehensive profiling of gene expression patterns within the intact tissue microenvironment. These techniques have become indispensable for studying complex spatiotemporal signaling events during critical developmental processes such as mouse gastrulation, where BMP, Nodal, and FGF pathways orchestrate cell fate decisions through precise coordination. While bulk and single-cell RNA sequencing have provided invaluable insights into cellular heterogeneity, they fundamentally lose the spatial context of cells within native tissue architecture [40]. This spatial context is particularly crucial for understanding morphogen gradients and signaling centers that pattern the embryo.
The emergence of spatial transcriptomics has coincided with advances in stem cell biology that enable the creation of in vitro models of early development. Gastruloids—three-dimensional stem cell aggregates that form an anterior-posterior axis and structures resembling the mammalian primitive streak and tailbud—provide a powerful experimental system for investigating symmetry breaking and pattern formation [23]. These systems capture essential features of in vivo development while offering unprecedented experimental accessibility for perturbing and measuring signaling dynamics.
Spatial Transcriptomic Technology Landscape
Spatial transcriptomic technologies have evolved along several technological trajectories, each with distinct advantages and limitations for profiling signaling components. These methods can be broadly classified into four categories based on their underlying technical principles [40].
Technology Classifications and Characteristics
In situ hybridization (ISH)-based technologies detect transcripts through hybridization of labeled complementary probes to mRNAs of interest within intact tissue sections. Single-molecule fluorescence in situ hybridization (smFISH) approaches provide high resolution at subcellular levels. Advanced multiplexed methods including sequential FISH (seqFISH) and multiplexed error-robust FISH (MERFISH) dramatically increase detection capacity through combinatorial barcoding strategies [40]. MERFISH combines combinatorial labeling with sophisticated encoding schemes to uniquely identify thousands of RNA species with error-robust detection [40]. seqFISH+ extends this approach with barcodes encoded across up to 60 channels, enabling comprehensive transcriptome profiling without requiring tissue clearance [40].
In situ sequencing (ISS)-based technologies directly sequence transcripts within tissue sections. While early ISS methods suffered from low capture efficiency, recent advances including STARmap and HybISS offer improved performance. STARmap provides higher detection efficiency than single-cell RNA sequencing and operates without reverse transcription, while HybISS increases throughput compared to conventional ISS approaches [40].
Next-generation sequencing (NGS)-based technologies capture RNA molecules from tissue sections onto spatially barcoded arrays for subsequent sequencing. The original Spatial Transcriptomics (ST) method and commercial implementations such as 10x Genomics Visium provide unbiased, whole-transcriptome coverage at resolutions of 55-100μm [40]. Higher-resolution methods including Slide-seq (10μm), HDST (2μm), and Seq-Scope (0.5-0.8μm) use DNA-barcoded beads to achieve cellular and subcellular resolution, while stereo-seq offers nanoscale resolution (500/715nm) across large detection areas [40].
Spatial information reconstruction technologies such as tomo-seq and STRP-seq computationally reconstruct spatial expression patterns from serially sectioned tissues without direct imaging, instead relying on algorithmic reconstruction of spatial patterns [40].
Table 1: Spatial Transcriptomic Technologies for Signaling Component Profiling
| Technology | Resolution | Detection Efficiency | Key Applications | Throughput |
|---|---|---|---|---|
| MERFISH | Subcellular | ~80% | Targeted signaling component profiling | High-plex (1000s of genes) |
| seqFISH+ | Subcellular | ~49% | Comprehensive signaling pathway analysis | High-plex (1000s of genes) |
| Visium | 55μm | Varies | Unbiased whole transcriptome | Whole transcriptome |
| Slide-seqV2 | 10μm | Higher than Slide-seq | Cellular-level spatial mapping | Whole transcriptome |
| DBiT-seq | 10μm | Varies | Multimodal spatial analysis | Whole transcriptome |
| STARmap | Subcellular | Higher than scRNA-seq | 3D signaling networks | Targeted |
Technology Selection Considerations
Choosing the appropriate spatial transcriptomic technology for profiling signaling components requires careful consideration of multiple parameters. The decision framework should prioritize spatial resolution needs (tissue, cellular, or subcellular), transcriptomic coverage (targeted vs. whole transcriptome), sensitivity for detecting low-abundance signaling components, multiplexing capacity for simultaneous pathway analysis, and compatibility with downstream assays [40]. For focused investigation of known signaling pathway components, targeted approaches such as MERFISH provide high sensitivity and quantification accuracy. For discovery-based studies of novel signaling interactions, unbiased NGS-based methods offer comprehensive coverage.
Spatial Mapping of Signaling Pathways in Mouse Gastrulation
Mouse gastrulation represents a paradigm for understanding how signaling pathways orchestrate complex morphogenetic events. During this critical developmental window, BMP, Nodal, and FGF pathways establish coordinated signaling gradients that pattern the embryonic axes and specify the three germ layers.
BMP Signaling Dynamics and Fate Determination
BMP signaling functions as a key morphogen with conserved roles in dorsoventral patterning. Recent studies using automated tracking of signaling histories in human pluripotent stem cells (hPSCs) revealed that the time-integral of BMP signaling—rather than instantaneous concentration—determines cell fate decisions [10]. This discovery fundamentally changes how we interpret morphogen gradients during gastrulation.
In micropatterned hPSC colonies that model human gastrulation, combined histories of BMP and Nodal signaling accurately predict cell fate patterns [10]. Live imaging of endogenously tagged SMAD4 (common mediator of BMP and Nodal signaling) and SMAD1 (BMP-specific) revealed distinct temporal signatures: sustained BMP signaling promotes amnion-like differentiation, transient BMP followed by Nodal activity correlates with primitive streak-like differentiation, and transient BMP without subsequent Nodal signaling maintains pluripotency [10]. This demonstrates how spatial transcriptomics can decode the logic of developmental signaling.
Nodal and Wnt Signaling in Axis Formation
The coordination between Nodal and Wnt signaling establishes the embryonic anterior-posterior axis. In mouse gastruloids, synthetic "signal-recording" gene circuits have traced how early heterogeneity in Nodal activity precedes and predicts subsequent Wnt patterning [23]. These circuits function as logical AND gates between a signaling pathway and a small molecule inducer, permanently labeling cells based on their signaling history.
This approach revealed that gastruloid polarization occurs through cell sorting mechanisms rather than reaction-diffusion systems alone. Initially patchy domains of Wnt activity become rearranged into a single posterior pole through cellular rearrangements, with Wnt-high cells sorting to define the elongating axis [23]. The origins of this Wnt heterogeneity can be traced to earlier Nodal signaling differences, illustrating a hierarchical relationship between these pathways.
Cross-Germ-Layer Signaling Networks
Advanced spatial transcriptomic approaches have enabled the reconstruction of full digital embryos during early mouse organogenesis (E7.5-E8.0). By profiling 285 serial sections from six embryos and developing the SEU-3D computational method, researchers established spatiotemporal transcriptome maps at single-cell resolution [41]. This revealed a "primordium determination zone" (PDZ) along the anterior embryonic-extraembryonic interface at E7.75, where coordinated signaling communications between germ layers contribute to cardiac primordium formation [41].
Space-informed gene-cell co-embedding approaches systematically characterized signaling networks across germ layers and cell types, revealing previously unappreciated signaling centers at tissue interfaces. These studies demonstrate how spatial transcriptomics moves beyond cataloging expression patterns to elucidating functional signaling circuits that span germ layer boundaries.
Table 2: Key Signaling Components in Mouse Gastrulation
| Signaling Pathway | Key Ligands/Receptors | Spatial Expression Pattern | Functional Role in Gastrulation |
|---|---|---|---|
| BMP | BMP4, BMPR1A/1B | Gradient from posterior to anterior | Dorsoventral patterning, amnion specification |
| Nodal | Nodal, ACVR1/2, SMAD2/3 | Anterior visceral endoderm, posterior | Anterior-posterior patterning, mesendoderm induction |
| Wnt | Wnt3, Frizzled, LRP5/6 | Primitive streak, posterior | Axis specification, mesoderm formation |
| FGF | FGF4, FGF8, FGFR1 | Primitive streak, emerging mesoderm | Mesoderm migration, epithelial-mesenchymal transition |
Experimental Frameworks and Methodologies
Signal-Recording Gene Circuits for Tracing Signaling Histories
Synthetic biology approaches now enable permanent recording of signaling pathway activities within developing systems. The Wnt-recorder circuit exemplifies this strategy [23]:
Circuit Design: A destabilized doxycycline-dependent transcription factor (rtTA) is expressed downstream of a TCF/LEF-responsive "sentinel enhancer." Combined Wnt activity and doxycycline presence trigger rtTA-dependent expression of a destabilized Cre recombinase, which mediates a permanent fluorescent switch from dsRed to GFP.
Implementation Protocol:
- Generate mouse embryonic stem cells (mESCs) harboring the Wnt-recorder circuit
- Culture in "2i+LIF" media to minimize pre-existing heterogeneity
- Aggregate cells to form gastruloids (400-1000 cells/aggregate)
- Pulse with CHIR-99021 (Wnt activator) for 24 hours (48-72 hours after aggregation)
- Administer doxycycline (100-200 ng/mL) for 1.5-3 hours during desired temporal window
- Fix samples at various timepoints or continue live imaging
Validation Experiments:
- Confirm specificity: No labeling with dox or CHIR alone
- Determine temporal resolution: Circuit reflects signaling changes within 6-hour windows
- Measure labeling efficiency: >68% with 1-hour dox pulse, near-complete with 3-hour pulse
This recording strategy enables fate mapping of signaling histories, linking early pathway activity to eventual cell position and fate decisions.
Live Imaging of Endogenous Signaling Activity
Direct visualization of signaling dynamics requires endogenously tagged pathway components [10]:
Cell Line Engineering:
- Generate knock-in alleles expressing GFP::SMAD4 or RFP::SMAD1 from endogenous loci
- Utilize CRISPR/Cas9-mediated homologous recombination
- Validate proper localization and signaling response
Imaging Setup:
- Culture micropatterned hPSC colonies or gastruloids in glass-bottom dishes
- Maintain at 37°C, 5% CO₂ during time-lapse imaging
- Acquire images at 20-60 minute intervals over 48-96 hours
- Quantify nuclear-to-cytoplasmic fluorescence ratios as signaling activity proxy
Data Processing:
- Segment individual nuclei across timepoints
- Track lineage relationships through mitosis
- Register cells to spatial coordinates within colony or gastruloid
- Compute radially averaged signaling patterns for population analysis
This approach revealed the zigzag temporal structure of SMAD4 signaling that predicts future cell fate boundaries.
Multimodal Spatial Data Integration
The SpatialData framework provides a unified approach for integrating diverse spatial omics datasets [42]:
Data Representation:
- Images: raster images (H&E, fluorescence)
- Labels: segmentation masks
- Points: molecular probes, single molecules
- Shapes: polygons (regions of interest, capture locations)
- Tables: molecular quantifications, annotations
Alignment Workflow:
- Define landmark points present across all datasets
- Compute transformation matrices to common coordinate system
- Validate alignment accuracy using known anatomical features
- Transfer spatial annotations between modalities
Cross-Modality Analysis:
- Aggregate single-cell data to spatial regions
- Transfer cell-type labels from scRNA-seq references
- Compare gene expression patterns across technologies
- Integrate histological features with molecular profiles
This framework enables robust comparison of signaling component localization across measurement platforms and spatial scales.
Computational and Data Analysis Approaches
Signaling History Analysis
Unbiased analysis of signaling dynamics requires specialized computational approaches [10]:
Principal Component Analysis (PCA) of Signaling Trajectories:
- Extract single-cell signaling histories as time-series vectors
- Perform PCA to identify dominant patterns of temporal variation
- Cluster cells based on PCA coordinates to identify qualitatively distinct signaling behaviors
- Map signaling clusters back to spatial positions
Time-Integral Modeling:
- Calculate cumulative signaling exposure as area under activity curve
- Test correlation between time-integral and differentiation outcomes
- Compare predictive power of integral models versus threshold models
- Validate through targeted perturbation of signaling duration versus intensity
This approach revealed that BMP signaling level and duration are interchangeable in fate specification, with both influencing differentiation only through their effect on the time-integral of signaling activity [10].
Digital Embryo Reconstruction
Three-dimensional reconstruction of full embryos enables comprehensive analysis of signaling networks [41]:
SEU-3D Method:
- Process serial tissue sections with spatial transcriptomics
- Align consecutive sections using anatomical landmarks
- Register single-cell transcriptomes to spatial coordinates
- Reconstruct three-dimensional expression patterns
- Model signaling gradients and cell-cell communication
Cross-Germ-Layer Analysis:
- Identify signaling sources and sinks across germ layers
- Infer ligand-receptor interactions between neighboring cell types
- Characterize signaling centers at tissue interfaces
- Map trajectory of cell migration in response to chemotactic signals
This systems-level approach revealed the primordium determination zone as a signaling center coordinating multi-germ-layer patterning.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Spatial Analysis of Signaling Pathways
| Reagent/Cell Line | Function/Application | Key Features | Example Use |
|---|---|---|---|
| Wnt-Recorder mESCs | Records Wnt signaling history | TCF/LEF-responsive; doxycycline-dependent | Fate mapping of Wnt-responsive cells [23] |
| GFP::SMAD4 mESCs | Live imaging of BMP/Nodal signaling | Endogenous tagging; proper localization | Quantitative signaling dynamics [10] |
| RFP::SMAD1 mESCs | Live imaging of BMP signaling | BMP-specific; endogenous tagging | Distinguishing BMP from Nodal activity [10] |
| 2i+LIF Media | Maintains ground pluripotency | Inhibits differentiation; minimizes heterogeneity | Pre-gastruloid culture [23] |
| CHIR-99021 | Wnt pathway activation | GSK3β inhibitor; uniform pathway activation | Gastruloid symmetry breaking [23] |
| SpatialData Framework | Multimodal data integration | Unified representation; cross-alignment | Integrating Xenium, Visium, H&E data [42] |
Visualizing Signaling Pathways and Experimental Workflows
Signaling Pathway Diagram
Signal Recording Workflow
Spatial Data Integration Framework
The precise functional analysis of signaling pathways—such as BMP, Nodal, and FGF—during mouse gastrulation requires techniques that can manipulate gene expression and protein activity with high spatiotemporal control. Among the most powerful methods for this purpose are in utero electroporation (IUE) for genetic manipulation and the application of small molecule inhibitors for acute protein inhibition. When used individually or in combination, these techniques enable researchers to dissect the complex roles of developmental signals in a manner that is often impossible with traditional transgenic models. IUE allows for the introduction of genetic constructs into specific tissue regions at precise developmental time points, while small molecule inhibitors provide temporal control over protein function. This guide details the methodology, integration, and application of these tools within the context of a research program focused on signaling pathways in early mouse development.
Technical Foundation of In Utero Electroporation
Core Principles and Mechanisms
In utero electroporation is a technique that enables the direct manipulation of gene expression in the developing embryonic brain in vivo. The fundamental process involves two key steps [43]:
- Electropermeabilization: A train of electrical pulses is applied, which temporarily disrupts the phospholipid bilayer of the cell membrane, creating pores.
- Electrotransfer: The electrical field also acts on the charged DNA plasmids present in the ventricle, driving them through the pores and into the cells lining the ventricular zone.
This method is particularly valuable for studying genes implicated in neurodevelopmental disorders, as it allows for the segregation of a gene's function in specific cell types and at particular developmental stages, thereby overcoming the limitations of full-knockout models which may have compensatory mechanisms or lethality [44] [45]. The technique's feasibility extends to behavioral examinations, allowing for the creation of animal models that recapitulate endophenotypes of complex psychiatric conditions [44].
Spatiotemporal Control of Gene Expression
A defining feature of IUE is the exquisite control it offers over where and when a gene is manipulated.
- Spatial Control: The site of DNA incorporation is determined by the position and orientation of the electrodes. By varying the electrode placement, researchers can target different brain regions, such as the hippocampus, prefrontal cortex, or striatum (Figure 1) [45]. Furthermore, the use of cell-type-specific promoters (e.g., Nestin for progenitor cells, DCX for post-mitotic migrating neurons) can restrict expression to particular neuronal subsets [44].
- Temporal Control: The developmental time point of electroporation determines the neuronal population affected. In the cerebral cortex, which develops in an "inside-out" manner, progenitor cells electroporated at E12.5, E13.5, or E14.5 give rise to pyramidal neurons predominantly in layers V/VI, IV, and II/III, respectively [44]. This allows researchers to interrogate the role of a gene in processes like proliferation, migration, or dendritic arborization by choosing the appropriate embryonic day.
Table 1: Targeting Specific Cell Populations via Electroporation Timing
| Electroporation Time Point | Primary Cell Population Targeted | Key Developmental Process |
|---|---|---|
| E12.5 | Cortical layers V/VI neurons | Early neurogenesis, migration |
| E13.5 | Cortical layer IV neurons | Mid neurogenesis, migration |
| E14.5 | Cortical layers II/III neurons | Late neurogenesis, migration |
| E18 (Rat) | Astrocytes | Gliogenesis |
For even more precise temporal control, inducible systems can be employed. These often use a Cre recombinase fused to a modified estrogen receptor (CAG-ERT2CreERT2), which is only activated upon injection of 4-hydroxytamoxifen (4-OHT). This allows the timing of gene expression to be controlled post-electroporation, segregating the effects of a genetic manipulation on a specific cellular process from secondary effects [44].
Advanced IUE Methodologies for Pathway Analysis
Combinatorial and Inducible Genetic Manipulation
The complexity of signaling pathways during gastrulation often requires the simultaneous manipulation of multiple genes. IUE is ideally suited for this, as multiple plasmids—carrying, for instance, overexpression constructs, short hairpin RNA (shRNA) for knockdown, or dominant-negative/constituitively-active forms of signaling components—can be co-electroporated [44]. This enables the functional testing of genetic interactions within common molecular pathways. For example, the role of BMP signaling in neural specification can be probed by co-electroporating a BMP inhibitor like Noggin or a constitutively active BMP receptor alongside regional fate markers.
Integration with Optogenetics
A powerful extension of IUE is its combination with optogenetics. By electroporating plasmids encoding light-sensitive opsins, researchers can gain millisecond-scale control over the activity of specific neuronal populations that were genetically targeted during development [43]. This approach allows for the functional mapping of neural circuits that arise from developmentally manipulated cells, linking early molecular interventions to later-stage functional outcomes in neural networks.
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents for IUE and Signaling Pathway Manipulation
| Reagent / Material | Function / Application | Technical Notes |
|---|---|---|
| Plasmid Vectors | Delivery of genetic material for overexpression, knockdown (shRNA), or reporter expression. | Can be episomal; for stable expression in proliferating cells, use transposon systems (e.g., PiggyBac, Sleeping Beauty) [43]. |
| 4-Hydroxytamoxifen (4-OHT) | Inducer for temporal control of gene expression in Cre-ERT2 systems. | Allows precise timing of genetic recombination and transgene activation post-electroporation [44]. |
| Small Molecule Inhibitors | Acute and reversible inhibition of specific signaling pathways (e.g., BMP, FGF, Nodal, Wnt). | Enables functional testing of pathway requirements in a time-resolved manner [15]. |
| Electroporator | Device generating square-wave electrical pulses for DNA delivery. | Requires precise control of voltage, pulse duration, and number of pulses [45]. |
| Fine Glass Micropipettes | For ultrasound-guided injection of DNA solution into the embryonic ventricle. | Essential for accurate and non-destructive delivery of constructs [45]. |
| Electrodes (paddle/wire) | Delivery of electrical current to the embryonic brain. | Electrode geometry and placement dictate the spatial specificity of transfection [45]. |
Signaling Pathways in Mouse Gastrulation: Context for Functional Studies
Mouse gastrulation is orchestrated by the tightly regulated activity of several key signaling pathways, including BMP, Wnt, Nodal, and FGF [12] [3]. These pathways interact to induce and pattern the primitive streak, guiding the cellular rearrangements that establish the three germ layers and the body plan. Our understanding of these events has been largely derived from genetic experiments in mouse embryos, though recent advances in embryo models like gastruloids are providing new insights into signaling dynamics [3].
The role of these pathways is complex and context-dependent. For instance, while Bmp signaling is classically known for its role in promoting epidermal fate in the ectoderm, studies in zebrafish have shown that in vegetal regions, Bmp activity is compatible with neural specification and instead promotes caudal character in the neural tissue [46]. Similarly, Fgf signaling has been shown to induce neural markers independently of Bmp antagonism in these regions [46]. In mouse embryonic stem cells (ESCs), BMP signaling can induce a totipotent state, but this role is constrained by its cross-activation of the FGF, Nodal, and WNT pathways [15]. This intricate cross-talk highlights the necessity of techniques like IUE and small molecule inhibition to disentangle the specific contributions of each pathway in a spatiotemporally defined manner.
Experimental Workflow: Integrating IUE and Small Molecule Inhibitors
The following diagram illustrates a generalized workflow for a functional experiment combining IUE and small molecule inhibitors to study signaling pathways in a developing mouse embryo.
Figure 1. Integrated experimental workflow for functional manipulation of signaling pathways. This workflow combines genetic manipulation via in utero electroporation with acute pharmacological inhibition to dissect pathway function.
Detailed Methodologies for Key Experiments
Protocol 1: In Utero Electroporation for Cortical Progenitors
- Timed Pregnancies: Establish timed matings, with the day of the vaginal plug designated as E0.5. Electroporation is typically performed between E12.5 and E15.5 for cortical studies.
- Surgical Preparation: Anesthetize the pregnant dam and perform a laparotomy to expose the uterine horns. Keep the embryos moist with sterile PBS throughout the procedure.
- DNA Injection: Using a finely pulled glass micropipette, inject ~1-2 µL of plasmid DNA solution (typically at 1-5 µg/µL, mixed with a fast green dye for visualization) into the lateral ventricles of the embryonic brain.
- Electroporation: Position paddle electrodes on either side of the injected embryo's head to target the desired brain region. Apply five 50 ms pulses of 30-40 V with 950 ms intervals using a square-wave electroporator.
- Post-operative Care: Return the uterus to the abdominal cavity, suture the dam, and allow the embryos to develop until the desired analysis stage [44] [45].
Protocol 2: Functional Interrogation of BMP Signaling
- Genetic Manipulation: Co-electroporate a plasmid of interest (e.g., a constitutively active BMP receptor, BMP shRNA, or a neutral control) with a fluorescent reporter (e.g., GFP) to mark transfected cells.
- Pharmacological Inhibition: At a specific time point post-electroporation (e.g., 24 hours later), administer a BMP pathway small molecule inhibitor (e.g., LDN-193189) to the pregnant dam via intraperitoneal injection or through the drinking water. The dose and duration depend on the inhibitor's pharmacokinetics and the desired window of inhibition.
- Analysis: Harvest embryos at a later developmental stage (e.g., E18.5) or postnatally. Analyze the phenotype using immunohistochemistry for neural markers (e.g., Sox3 for prospective neural tissue, p63 for epidermal fate), and assess neuronal migration, layer formation, and axon guidance [46] [15]. The use of both genetic and pharmacological tools allows for the validation and precise timing of BMP pathway manipulation.
Quantitative Data and Analysis
Table 3: Quantitative Parameters for IUE and Pathway Manipulation
| Parameter | Typical Range / Value | Significance / Impact |
|---|---|---|
| DNA Concentration | 1 - 5 µg/µL | Lower concentrations may yield weak expression; higher concentrations can be toxic [45]. |
| Electroporation Voltage | 30 - 40 V (mice) | Optimized to balance transfection efficiency and embryo survival [45]. |
| Pulse Duration | 50 ms | Sufficient for membrane permeabilization and DNA transfer [43]. |
| BMP Inhibitor (LDN-193189) Dose | 3 - 10 mg/kg (in vivo) | Effective for systemic inhibition of BMP type I receptors (ALK2/3) [15]. |
| Contrast Ratio for Visualization | ≥ 4.5:1 (small text) ≥ 3:1 (large text) | Critical for accessibility of diagrams and data presentation, ensuring information is perceivable by all [47] [48]. |
In utero electroporation and small molecule inhibitors represent a formidable pair of technologies for the functional dissection of signaling pathways in developing systems. The ability to spatially and temporally control genetic manipulations with IUE, combined with the acute temporal precision of small molecule inhibitors, provides a powerful means to interrogate the complex interactions of pathways like BMP, FGF, and Nodal during critical developmental windows such as gastrulation and neural development. By applying the detailed methodologies and reagents outlined in this guide, researchers can design robust experiments to causally link molecular pathways to cellular, circuit, and behavioral phenotypes, thereby advancing our understanding of both normal development and the etiology of disease.
Gastrulation is a fundamental developmental process in mouse embryos whereby cells exit a state of pluripotency and concomitantly acquire and pattern distinct cell fates [49]. This complex transformation is primarily driven by the convergence of four key signaling pathways: WNT, BMP, Nodal, and FGF [49] [3]. These morphogen signals are tightly spatially and temporally controlled within the embryo, resulting in regional and stage-specific signaling environments that dictate the eventual fate of each cell according to its position and developmental timing [49]. The combination, level, and duration of signals a cell is exposed to determines whether it will contribute to ectodermal, mesodermal, or endodermal lineages [49].
Understanding the dynamics of these signaling pathways is crucial for developmental biology research, particularly because abnormal phosphorylation states and signaling disruptions are implicated in many human diseases, including cancer and developmental disorders [50]. The intricate interactions between these pathways make them difficult to disentangle in vivo, necessitating the development of sophisticated analytical tools including reporter assays and phospho-specific antibodies [49] [23]. Recent advances in stem cell models, particularly gastruloids, have provided powerful in vitro systems that undergo organized cellular differentiation and patterning, serving as simplified, deconstructed, and scalable models of gastrulation that are revealing new insights into signaling regulation [49] [3].
Reporter Assays for Monitoring Pathway Activity
Engineering and Applications of Signal-Recorder Circuits
Reporter assays represent a powerful approach for monitoring the activity of signaling pathways in real-time within living cells and tissues. Recent innovations have led to the development of sophisticated "signal-recording" gene circuits that can permanently record transient signaling events, enabling researchers to trace the evolution of signaling patterns over time [23]. These circuits function as logical AND gates that require both a candidate signaling pathway and a user-supplied small molecule to produce a permanent, heritable fluorescent signal [23].
The fundamental design of these circuits typically involves expressing a destabilized doxycycline-dependent transcription factor (rtTA) downstream of a 'sentinel enhancer' that responds to a specific pathway of interest (e.g., TCF/LEF for Wnt signaling) [23]. The combined presence of both signaling activity and doxycycline triggers activation of an rtTA-dependent promoter (PTetON) to drive expression of a destabilized Cre recombinase, which in turn results in a permanent change in fluorescent protein expression (e.g., from dsRed to GFP) [23]. Because these recordings are inherited by all progeny of originally labeled cells, they capture differences in proliferation and cell migration between the initial recording and final read-out measurement [23].
In practice, these reporter systems have demonstrated remarkable sensitivity and specificity. For Wnt-responsive recorders, no detectable labeling occurs when cells are incubated with either doxycycline or Wnt activator (CHIR) alone, requiring both inputs for signal generation [23]. Labeling efficiency remains exceptionally low (<0.1%) over multiple passages in Wnt-activating conditions without doxycycline, demonstrating minimal leakiness [23]. These systems can detect signaling events within relatively brief time windows, with efficient labeling achievable with doxycycline pulses as short as 1-3 hours [23]. There is typically a 3-6 hour delay before the labeling circuit fully reflects changes in pathway activity, attributable to the time required for rtTA expression or turnover [23].
Experimental Workflow for Signal Recording
The experimental implementation of signaling recorder circuits follows a structured workflow that begins with the aggregation of mouse embryonic stem cells (mESCs) into gastruloids [23]. These aggregates are typically treated with the Wnt pathway activator CHIR-99021 between 48 and 72 hours after aggregation to initiate symmetry breaking and patterning events [23]. During critical time windows of interest, researchers administer brief pulses of doxycycline (100-200 ng/mL for 1.5-3 hours) to activate the recording circuit specifically in cells experiencing signaling activity [23]. The combined presence of both the pathway activity and doxycycline triggers permanent fluorescent protein expression in responding cells and all their progeny [23]. Finally, researchers can perform lineage tracing and quantitative analysis of the labeled populations to understand how early signaling states correlate with eventual cell fates and positions along the anterior-posterior axis [23].
Key Insights from Reporter Studies in Gastruloids
Application of these reporter assays to mouse gastruloids has revealed fundamental insights into the self-organization of embryonic structures. Studies have demonstrated that gastruloids progress through distinct phases of Wnt signaling: initially uniform low activity at 48 hours after aggregation (haa), shifting to uniformly high activity by 72 haa following CHIR treatment, then developing bimodal distributions with Wnt-high and Wnt-low subpopulations, and finally establishing a single coherent domain of Wnt activity that marks the elongating posterior by 108 haa [23]. This polarization process begins with the emergence of patchy, locally ordered domains of Wnt activity around 96 haa that subsequently resolve into a single posterior pole [23].
These reporter studies have provided crucial evidence distinguishing between competing models of symmetry breaking. The observation that early Wnt domains predict cells' ultimate positions along the anterior-posterior axis, even before global Wnt patterns become polarized, supports a cell-sorting mechanism where an initial mixture of Wnt-high and Wnt-low cells rearranges to achieve polarization, rather than a reaction-diffusion mechanism where feedback loops gradually refine cells' signaling states [23]. Furthermore, these approaches have enabled researchers to trace the emergence of Wnt heterogeneity to even earlier heterogeneity in Nodal and BMP signaling, revealing a hierarchical organization of patterning cues [23].
Phospho-Specific Antibodies for Signaling Analysis
Development and Validation of Phospho-Specific Antibodies
Phospho-specific antibodies are vital tools for studying cell signaling pathways, enabling researchers to detect and quantify specific phosphorylation events that regulate protein function and activity [51] [52]. These antibodies are typically generated by immunizing host animals with short, chemically synthesized peptides containing a phosphorylated residue (serine, threonine, or tyrosine) conjugated to an immunogenic carrier protein such as Keyhole Limpet Hemocyanin (KLH) [52]. The resulting antisera undergo rigorous purification processes, often using affinity chromatography with phospho-peptide and non-phosphopeptide columns, to select for antibodies that specifically recognize the phosphorylated epitope while minimizing cross-reactivity with non-phosphorylated forms [52].
The specificity validation of phospho-specific antibodies represents a critical step in ensuring reliable experimental results. Recent approaches have introduced quantitative parameters to characterize antibody specificity more rigorously. One such parameter, termed Φ (phi), measures the fraction of non-specific signal in antibody binding and is determined using flow cytometry-based assays in engineered cell lines [53]. In this system, cells expressing either wild-type tau-EGFP or tau-iRFP with alanine mutations at phosphorylation sites are mixed and labeled with the phospho-specific antibody of interest [53]. For a perfectly specific antibody, binding should only occur to the wild-type tau-EGFP cells, with any binding to the alanine mutant tau-iRFP cells representing non-specific signal [53]. Applications of this validation method to commonly used phospho-tau antibodies revealed that many high-quality reagents (including AT8, AT180, PHF-6, TG-3, and PHF-1) exhibit Φ values near 1, indicating nearly perfect specificity, while others (such as AT270) showed detectable non-specific binding with Φ values around 0.8 [53].
Detection Methods Using Phospho-Specific Antibodies
Phospho-specific antibodies enable multiple detection modalities, each with distinct advantages and applications. The Western blot remains the most common method for assessing protein phosphorylation, combining SDS-PAGE separation with subsequent immunoblotting using phospho-specific antibodies [51]. This approach provides information about protein molecular weight and allows simultaneous detection of total protein levels using antibodies that recognize the target regardless of phosphorylation state [51]. Automated capillary-based Western systems (Simple Western) offer enhanced sensitivity, requiring only 3 μL of sample while providing multiplex detection capabilities for low-abundance phosphorylated and total protein isoforms [51]. These systems can process up to 96 samples in a single automated run and can separate phosphorylated protein isoforms using capillary isoelectric focusing (cIEF) [51].
Enzyme-linked immunosorbent assays (ELISAs) provide more quantitative measurements of protein phosphorylation and are particularly valuable for studies modulating kinase activity and function [51]. In the sandwich ELISA format, a capture antibody specific for the target protein (phosphorylation-independent) immobilizes the antigen, while a phospho-specific detection antibody enables quantification of the phosphorylated form [51]. ELISAs offer greater sensitivity and throughput than traditional Western blotting, with the microplate format facilitating the analysis of multiple samples in parallel [51]. More recently, multiplex immunoassays have been developed that enable simultaneous detection and quantitation of both phosphorylated and total forms of multiple proteins in a single well, providing comprehensive signaling pathway analysis [50].
For single-cell analysis, intracellular flow cytometry and immunocytochemistry/immunohistochemistry (ICC/IHC) represent powerful applications of phospho-specific antibodies [51]. Flow cytometry allows rapid, quantitative analysis of phosphorylation events at single-cell resolution within heterogeneous populations, enabling researchers to detect signaling differences in rare cell subsets without physical separation [51]. ICC/IHC provides spatial context for phosphorylation events within tissue architecture or subcellular compartments, although with more limited quantification capabilities [51].
Comparative Analysis of Detection Methods
Performance Characteristics and Applications
Table 1: Comparison of Major Protein Phosphorylation Detection Methods
| Method | Best Application | Sample Type | Quantification | Multiplexing | Key Advantages |
|---|---|---|---|---|---|
| Western Blot | Validation, confirmation, visual identification | Cell/tissue lysates (denatured) | Semi-quantitative | Limited (up to 4-plex with fluorescence) | Protein size information, widely accessible |
| Simple Western | Automated Western analysis, low sample volume | Cell/tissue lysates (denatured) | Quantitative with standards | Moderate (multiplex detection available) | High sensitivity, automated, minimal sample required |
| ELISA | Single-target analysis with high specificity | Various (lysates, serum, plasma, supernatants) | Quantitative with standard curve | No (single analyte) | Excellent quantification, high throughput, sensitive |
| Multiplex Immunoassays | Profiling multiple targets simultaneously | Various (native conditions) | Quantitative with standard curve | High (up to 80 analytes) | Comprehensive pathway analysis, sample efficient |
| Intracellular Flow Cytometry | Single-cell analysis in heterogeneous populations | Single cell suspensions | Quantitative | Moderate (limited by fluorochrome spectra) | Single-cell resolution, rapid analysis |
| ICC/IHC | Spatial context within tissues/cells | Fixed cells/tissues | Semi-quantitative | Moderate (limited by antibody host species) | Spatial information, morphological context |
Technical Considerations for Method Selection
The choice between detection methods depends on multiple factors including the specific research question, required throughput, sample availability, and necessary quantification rigor. Western blotting remains the most accessible technique for most laboratories and provides important information about protein molecular weight, which can help confirm target identity and detect potential proteolytic fragments or alternative isoforms [51]. However, Western blotting is generally considered semi-quantitative at best and suffers from relatively low throughput compared to plate-based assays [51]. The recent development of automated capillary Western systems (Simple Western) addresses many of these limitations by providing true quantitation, higher sensitivity, and greatly increased throughput while maintaining the protein size information and denaturing conditions of traditional Western blotting [51].
For studies requiring precise quantitation of phosphorylation levels or higher throughput analysis, ELISA formats offer significant advantages [51] [50]. Phospho-specific ELISAs are typically more sensitive than Western blotting, requiring 2-4 times less lysate for equivalent detection, and provide direct quantitative measurements through standard curves [50]. The sandwich ELISA format, using a capture antibody against the total protein and a detection antibody against the phosphorylated form, ensures high specificity through the requirement of two independent binding events [50]. These characteristics make ELISAs particularly valuable for dose-response studies, time-course experiments, and screening applications where precise quantitation and higher throughput are essential [51] [50].
When analyzing complex biological systems with limited sample availability, multiplex immunoassays provide unparalleled efficiency by enabling simultaneous measurement of multiple phosphorylated targets alongside their total protein counterparts [50]. These bead-based assays can quantify up to 80 analytes from a single small volume sample, making them ideal for comprehensive signaling pathway analysis or for working with precious clinical specimens [50]. For heterogeneous cell populations, intracellular flow cytometry offers unique insights by enabling phosphorylation analysis at single-cell resolution, allowing researchers to identify signaling differences between subpopulations that would be masked in bulk measurements [51].
Research Reagent Solutions for Gastrulation Signaling Studies
Table 2: Essential Research Reagents for Signaling Pathway Analysis
| Reagent Category | Specific Examples | Key Applications | Technical Considerations |
|---|---|---|---|
| Phospho-Specific Antibodies | Anti-phospho-Ser/Thr/Tyr antibodies; AT8, AT180, PHF-1 for tau phosphorylation | Western blot, ICC/IHC, flow cytometry | Require rigorous validation; check Φ values for specificity assessment |
| Signal Recording Systems | TCF/LEF-responsive reporters; doxycycline-inducible circuits | Live-cell imaging, lineage tracing, fate mapping | Temporal resolution ~3-6 hours; optimize doxycycline concentration and pulse duration |
| Pathway Modulators | CHIR-99021 (Wnt activator); DMH1 (BMP inhibitor) | Pathway perturbation studies, dose-response experiments | Determine optimal concentrations and treatment windows for specific model systems |
| ELISA Kits | Phospho-EGF R ELISA; STAT3 (Phospho) ELISA; CREB (Total/Phospho) ELISA | Quantitative phosphorylation assessment, high-throughput screening | Choose between standard sandwich format vs. rapid InstantOne kits based on throughput needs |
| Multiplex Immunoassays | ProcartaPlex Signaling panels; Dual Reporter assays | Comprehensive pathway analysis, limited sample availability | Validate antibody cross-reactivity; ensure compatibility with detection instrumentation |
| Cell Line Engineering Tools | CRISPR/Cas9 systems; lentiviral transduction | Endogenous tagging, reporter cell line generation | Consider knock-in efficiency and selection strategies; verify normal signaling function |
Integrated Experimental Approaches in Gastrulation Research
Combining Techniques for Comprehensive Signaling Analysis
The most powerful insights into signaling pathway activity during mouse gastrulation often come from integrated approaches that combine multiple analytical techniques. Reporter assays provide dynamic, temporal information about pathway activity in living systems, while phospho-specific antibodies offer snapshot quantification of specific molecular events with cellular resolution [23] [51]. The combination of these approaches enables researchers to correlate pathway activity with downstream phosphorylation events, creating a more complete understanding of signaling networks.
Recent studies of gastruloid development exemplify this integrated approach. By using Wnt-responsive reporter circuits, researchers demonstrated that gastruloids break symmetry through a cell sorting mechanism wherein initially patchy domains of Wnt activity resolve into a single posterior pole [23]. Complementary phospho-specific antibody studies could then quantify the activation states of downstream signaling components, verifying the functional consequences of these Wnt activity patterns [51] [50]. Similarly, the identification of pre-existing Nodal and BMP signaling heterogeneity as upstream regulators of Wnt patterning emerged from combined reporter and antibody-based approaches [23].
Protocol for Integrated Signaling Analysis in Gastruloids
A representative integrated protocol for analyzing signaling pathways in mouse gastruloids begins with the aggregation of mouse embryonic stem cells (mESCs) in low-cell-adhesion 96-well plates, with approximately 300-500 cells per aggregate in N2B27 media [23]. For reporter assays, utilize mESCs harboring Wnt-responsive (TCF/LEF) or other pathway-specific reporter constructs, treating aggregates with CHIR-99021 (3 μM) between 48-72 hours after aggregation to induce patterning [23]. To record signaling activity during critical windows, administer doxycycline pulses (100-200 ng/mL for 1.5-3 hours) at specific timepoints, then continue culture without doxycycline to allow permanent fluorescent labeling of cells with active signaling [23].
For parallel phospho-specific analysis, harvest gastruloids at multiple timepoints for fixed-sample analysis. For Western blotting, lyse gastruloids in RIPA buffer supplemented with phosphatase and protease inhibitors, separate 10-30 μg protein by SDS-PAGE, transfer to PVDF membranes, and probe with phospho-specific antibodies against targets of interest (e.g., phospho-SMAD1/5/8 for BMP signaling) followed by appropriate secondary antibodies [51]. Always include antibodies against total protein forms and loading controls to normalize phosphorylation levels [51]. For quantitative comparisons, use phospho-specific ELISAs according to manufacturer protocols, typically involving sample addition to antibody-coated plates, incubation with detection antibodies, and colorimetric or fluorometric measurement [50]. For single-cell analysis, dissociate gastruloids to single cells and analyze by intracellular flow cytometry using phospho-specific antibodies, ensuring proper fixation and permeabilization to maintain antibody accessibility while preserving cellular integrity [51].
Finally, integrate data from reporter assays, phospho-specific detection methods, and morphological assessments to build a comprehensive understanding of signaling dynamics during gastrulation events. Correlate early signaling activity (from reporter recordings) with subsequent phosphorylation events and ultimate cell fate decisions to establish causal relationships within the signaling network [23].
The analysis of signaling pathway activity during mouse gastrulation relies on two complementary technological approaches: reporter assays that provide dynamic, temporal information in living systems, and phospho-specific antibodies that enable precise molecular quantification of phosphorylation events. The continuing refinement of these tools, including the development of more sensitive recording circuits and increasingly specific antibodies, promises to reveal new insights into the complex signaling networks that orchestrate embryonic development. As these techniques become more sophisticated and accessible, they will undoubtedly enhance our understanding of both normal development and disease processes rooted in signaling pathway dysregulation.
Computational Modeling of Signaling Dynamics and Network Interactions
Embryonic development is orchestrated by complex signaling networks that direct cellular differentiation and tissue patterning. In mammalian gastrulation, secreted signaling molecules—including Bone Morphogenetic Protein (BMP), Wnt, Nodal, and Fibroblast Growth Factor (FGF)—coordinate to induce and pattern the primitive streak, initiating the cellular rearrangements that establish the basic body plan [12] [3]. Our understanding of how these pathways specify and organize germ layers in three dimensions stems largely from decades of genetic experimentation in mouse embryos [12]. However, the precise spatiotemporal sequence of signaling events remains incompletely understood due to the molecular and cellular complexity of these early developmental processes and a historical lack of tractable models for real-time visualization [12] [3].
The emergence of in vitro embryo models, particularly gastruloids derived from embryonic stem (ES) cells, has begun to illuminate the dynamics of signaling during primitive streak formation [12] [3]. These models provide a powerful platform for perturbing signaling pathways and observing outcomes in a controlled environment. Concurrently, advances in computational methods are enabling researchers to characterize and learn from the inherent heterogeneity in single-cell signaling data, moving beyond population averages to understand cell-to-cell variability and its functional consequences [54]. This technical guide integrates these experimental and computational frontiers, providing a framework for studying signaling network interactions within the context of mouse gastrulation.
Computational Approaches for Analyzing Signaling Dynamics
The Challenge of Heterogeneity in Signaling Data
A fundamental feature of cell signaling is heterogeneity: genetically identical cells, exposed to identical stimuli, often exhibit markedly different responses [54]. This heterogeneity arises from both intrinsic noise (random fluctuations in biochemical reactions involving low-abundance molecules) and extrinsic noise (preexisting differences in cell state, such as cell-cycle stage or pathway molecule concentrations) [54]. Disentangling these sources is crucial, as they have different implications for cellular control mechanisms and potential therapeutic interventions [54].
Data-Driven Computational Methods
Computational techniques are essential for extracting meaning from complex, dynamic single-cell signaling data obtained via live-cell microscopy and fluorescent reporters [54]. The following table summarizes key computational methods for analyzing such data:
Table 1: Computational Methods for Analyzing Heterogeneous Single-Cell Signaling Data
| Method | Core Function | Key Application in Signaling Studies | Benefits | Shortcomings |
|---|---|---|---|---|
| Clustering | Identifies families of distinct signaling trajectories by calculating distances between single-cell time series [54]. | Classifying ERK activation dynamics (e.g., transient vs. sustained) induced by different growth factors [54]. | Aggregates data based on real cell behaviors; simplifies analysis and presentation. | Can be inappropriate for continuous, non-discrete response patterns [54]. |
| Dimensionality Reduction (e.g., FPCA) | Projects high-dimensional data into a lower-dimensional space defined by primary axes of variation [54]. | Visualizing the major patterns of heterogeneity within a cell population's signaling dynamics [54]. | Enables easy visualization and identification of patterns in complex datasets. | The primary axes of variation may not correspond to physiologically meaningful processes [54]. |
| Mechanistic Modeling | Constructs mathematical models (e.g., ODEs) of signaling networks to simulate and understand underlying molecular interactions. | Understanding how pre-existing differences in cell state lead to heterogeneous pathway activation [54]. | Provides a causal, systems-level understanding of network behavior and regulation. | Model construction and parameterization can be complex and require extensive data. |
| Information Theory | Quantifies the mutual information between an input stimulus and a cellular response [54]. | Measuring the fidelity of information transmission through a specific signaling pathway. | Provides a model-free measure of signaling efficiency and channel capacity. | Does not provide mechanistic insights into the network structure. |
These methods move beyond simple summary statistics, which can obscure true single-cell behaviors—for example, by blurring a binary (ON/OFF) response into an intermediate value that represents no actual cell [54].
Experimental Foundation: Signaling in Mouse Gastrulation and Models
Core Signaling Pathways and Their Roles
During murine gastrulation, signaling gradients pattern the embryo. The extraembryonic ectoderm (ExE) expresses BMP4, establishing a proximal-to-distal gradient of phosphorylated SMAD1/5 [9]. This gradient promotes the differentiation of proximal mesoderm subtypes, such as extraembryonic mesoderm and blood [9]. Complementary gradients of Wnt and Nodal signaling, established by posterior ligand expression and anterior antagonists from the anterior visceral endoderm (AVE), pattern the distal mesoderm and endoderm, respectively [9].
Antagonistic Signaling Interactions: FGF and BMP
A critical interaction occurs between FGF and BMP signaling. Single-cell RNA sequencing analyses of differentiating stem cell populations reveal that FGF signaling operates within a positive autoregulatory loop (boosting expression of endogenous Fgf genes) while simultaneously repressing the expression of Bmp ligands [9]. This population-level antagonism helps specify coherent cohorts of distal mesoderm cells, spatially segregating them from BMP-dependent proximal cell types [9].
The following table quantifies the concentration-dependent effects of BMP and FGF signaling on key mesodermal markers, as observed in EpiSC differentiation models:
Table 2: Concentration-Dependent Effects of BMP and FGF Signaling on Mesoderm Markers in EpiSC Differentiation
| Signaling Factor | Concentration Range | Target Marker | Marker Expression Trend | Interpreted Fate Specification |
|---|---|---|---|---|
| BMP4 [9] | 16 - 32 ng/ml | Hand1, Gata6 (proximal markers) | Highest Expression | Proximal Mesoderm (e.g., extraembryonic, heart) |
| BMP4 [9] | ~4 ng/ml | T/Bra (pan-mesoderm), Tbx6, Msgn1 (distal/paraxial markers) | Peak Expression | Paraxial & Distal Mesoderm |
| BMP4 [9] | <4 ng/ml | Foxa2, Shh (most distal/axial markers) | Peak Expression | Axial Mesoderm & Endoderm |
| FGF2/FGF4 [9] | Titration in presence of BMP | T/Bra (pan-mesoderm) | Promotes Expression | General Mesoderm Differentiation |
Detailed Experimental Protocol: EpiSC Differentiation for Mesoderm Patterning Studies
This protocol outlines the key methodology for investigating BMP and FGF signaling in mesoderm differentiation, derived from recent studies [9].
Cell Culture and Initial Maintenance
- Cell Line: Mouse Epiblast Stem Cells (EpiSCs).
- Maintenance Medium (FAX): Culture EpiSCs in N2B27 basal medium supplemented with:
- ActivinA: To maintain pluripotency.
- FGF2: To support EpiSC state.
- XAV939 (Wnt inhibitor): To suppress Wnt-induced heterogeneity and maintain a homogeneous starting population [9].
- Validation: Confirm a homogeneous starting state by immunostaining for high NANOG (positive) and absence of the pan-mesodermal marker T/BRA (negative) [9].
Mesoderm Differentiation and Signaling Perturbation
- Differentiation Trigger: Replace the maintenance medium (ActivinA and XAV939) with differentiation medium containing:
- CHIR99021 (1 µM): A Wnt agonist to initiate mesoderm differentiation [9].
- BMP4: Use a concentration series (e.g., 0 ng/ml to 64 ng/ml) to probe its concentration-dependent effects on proximal-distal patterning [9].
- FGF2 or FGF4: Use a titration series in the presence of a intermediate BMP4 concentration (e.g., 8 ng/ml) to map FGF-dependent mesodermal lineages and its interaction with BMP [9].
- Time Course: Differentiate cells for approximately 3 days before analysis.
Readouts and Analysis
- Molecular Phenotyping:
- qPCR: Analyze expression of a panel of region-specific markers (e.g., Hand1, Gata6 for proximal; Tbx6, Msgn1 for distal; Foxa2, Shh for axial) [9].
- Single-Cell RNA Sequencing: Resolve heterogeneous cell populations and identify novel FGF-dependent and -independent lineages at high resolution [9].
- In-Cell Western: Quantify protein levels of key markers (e.g., HAND1, GATA6) across different signaling factor concentrations [9].
- Live-Cell Imaging: Utilize fluorescent reporter cell lines (e.g., T/Bra:mCherry) to monitor signaling dynamics and differentiation in real time [54] [9].
Diagram 1: EpiSC differentiation and signaling perturbation workflow.
Integrated Signaling Network Analysis and Computational Modeling
The integration of experimental data from embryo models with computational approaches is key to building a predictive understanding of gastrulation. Gastruloids, which self-organize and recapitulate aspects of embryonic patterning, serve as a powerful validation platform for computational models [12] [3]. Computational models can simulate the dynamics of the signaling network, testing hypotheses about how interactions like the FGF-BMP antagonism emerge from the underlying gene regulatory network [12].
The signaling network governing mesoderm patterning can be conceptualized as a system of interconnected pathways. The following diagram summarizes the core components and their known interactions, based on the discussed research:
Diagram 2: Core signaling network in murine gastrulation patterning.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Studying Signaling in Gastrulation Models
| Reagent / Tool | Category | Primary Function in Research |
|---|---|---|
| EpiSCs (Epiblast Stem Cells) [9] | Cell Model | Pluripotent cell population representing the post-implantation epiblast; used for 2D differentiation studies and signaling perturbation. |
| Gastruloids [12] [3] | Embryo Model | 3D aggregates of ES cells that self-organize and mimic aspects of embryonic gastrulation; used for studying signaling dynamics in a tissue context. |
| BMP4 [9] | Signaling Factor | Recombinant protein used to activate BMP signaling; titrated to specify proximal mesoderm fates in differentiation protocols. |
| FGF2 / FGF4 [9] | Signaling Factor | Recombinant proteins used to activate FGF signaling; required for efficient mesoderm differentiation and distal fate specification. |
| CHIR99021 [9] | Small Molecule Agonist | GSK-3 inhibitor that activates Wnt/β-catenin signaling; used to trigger mesoderm differentiation. |
| XAV939 [9] | Small Molecule Inhibitor | Tankyrase inhibitor that suppresses Wnt/β-catenin signaling; used to maintain a homogeneous EpiSC population. |
| T/Bra:mCherry Reporter [9] | Reporter Cell Line | Genetically engineered cell line where mCherry fluorescence reports the expression of the pan-mesodermal marker T/Brachyury; enables live imaging of mesoderm differentiation. |
| Single-Cell RNA Sequencing [54] [9] | Analytical Tool | High-resolution transcriptomics used to map heterogeneous cell populations, identify novel lineages, and infer signaling states from ligand/receptor expression. |
Experimental Challenges: Pathway Modulation and Data Interpretation
Overcoming Pathway Redundancy and Compensation Mechanisms
The establishment of the mammalian body plan during gastrulation is orchestrated by a complex interplay of key signaling pathways, primarily Bone Morphogenetic Protein (BMP), Nodal, and Fibroblast Growth Factor (FGF). These pathways form an intricate gene regulatory network that directs cell fate specification, patterning, and morphogenesis. A significant challenge in developmental biology and therapeutic intervention is the inherent redundancy and compensatory mechanisms within these signaling networks, where inhibition of one pathway can be offset by the modulation of others. In mouse gastrulation, BMP, Nodal, and FGF signaling interact through cross-activation, mutual antagonism, and feedback loops, creating a robust system that ensures developmental precision but complicates experimental and therapeutic manipulation. This whitepaper provides an in-depth technical analysis of these compensatory mechanisms and presents validated experimental strategies to overcome them, providing a critical resource for researchers and drug development professionals working in this field.
Core Signaling Pathways and Their Interactions
The BMP, Nodal, and FGF pathways each play distinct yet interconnected roles during mouse gastrulation. BMP signaling, characterized by a gradient of activity, promotes the differentiation of proximal mesoderm subtypes and is crucial for primitive streak induction [3] [9]. Nodal, a TGF-β family member, is essential for mesendoderm formation and establishes the anterior-posterior axis [3]. FGF signaling drives epithelial-to-mesenchymal transition, promotes the specification of distal mesoderm lineages such as paraxial mesoderm, and exhibits a strong antagonistic relationship with BMP signaling [9] [55]. The regulatory logic between these pathways is characterized by two dominant features: (1) antagonistic relationships, where FGF signaling represses BMP ligand expression and promotes its own expression through positive autoregulation; and (2) cross-activation mechanisms, where BMP signaling can initiate FGF, Nodal, and Wnt pathways, thereby constraining its own specific effects [9] [15].
Visualizing the Signaling Network
The following diagram illustrates the core interactions and compensatory mechanisms between BMP, FGF, and Nodal pathways during mouse gastrulation:
Figure 1: Signaling pathway interactions and compensation mechanisms in mouse gastrulation. BMP promotes proximal mesoderm formation while FGF specifies distal mesoderm lineages. Antagonistic relationships (dashed arrows) and cross-activations create inherent redundancy.
Quantitative Analysis of Pathway Interactions
BMP and FGF Dose-Dependent Effects on Mesoderm Specification
Quantitative studies in epiblast stem cell (EpiSC) differentiation systems reveal how BMP and FGF signaling concentrations direct mesodermal subtype specification. The data below summarize concentration-dependent marker expression in response to BMP4 and FGF stimulation:
Table 1: Concentration-dependent effects of BMP4 on mesodermal marker expression
| BMP4 Concentration (ng/mL) | Peak Marker Expression | Expression Level | Biological Interpretation |
|---|---|---|---|
| 0-2 ng/mL | Foxa2, Shh | High at lower concentrations | Distal/anterior patterning |
| ~4 ng/mL | T/Bra, Tbx6, Msgn1 | Peak expression | Pan-mesoderm and distal mesoderm |
| 16-32 ng/mL | Hand1, Gata6 | High expression | Proximal mesoderm specification |
| >32 ng/mL | HAND1 (protein) | Continues increasing | Extraembryonic mesoderm commitment |
Source: Adapted from single-cell analysis of EpiSC differentiation [9]
Table 2: FGF-mediated opposition of BMP signaling and autoregulation
| Experimental Condition | BMP Target Response | FGF Target Response | Network Effect |
|---|---|---|---|
| Low FGF stimulation (≤10 ng/mL FGF2) | Enhanced BMP signaling | Minimal autoregulation | Proximal mesoderm bias |
| High FGF stimulation (≥50 ng/mL FGF2) | Repressed Bmp ligand genes | Endogenous Fgf gene upregulation | Distal mesoderm commitment |
| FGF pathway inhibition | BMP signaling expansion | Loss of positive autoregulation | Disrupted community effect |
Source: Analysis of FGF-BMP antagonism in mesoderm patterning [9]
Temporal Dynamics of BMP/Nodal Ratios
The timing of pathway activation creates a developmental window during which signaling ratios determine tissue-specific morphogenetic programs. Research in zebrafish models demonstrates that the BMP/Nodal ratio during early gastrulation stages directs convergence and extension movements in specific tissues:
Table 3: Temporal effects of BMP/Nodal signaling ratios on tissue morphogenesis
| Developmental Window | BMP/Nodal Ratio | Morphogenetic Outcome | Molecular Mechanism |
|---|---|---|---|
| Early gastrulation | Low BMP/Nodal | Mesoderm-driven convergent extension | Nodal-induced mesoderm specification |
| Early-to-mid gastrulation | High BMP/Nodal | Neuroectoderm-driven extension | BMP-mediated repression of mesoderm |
| Critical temporal window | Dynamic ratio shift | Tissue-specific morphogenesis | Sequential activation of distinct gene programs |
Source: Zebrafish explant studies of BMP/Nodal temporal dynamics [22]
Experimental Strategies to Overcome Compensation
Simultaneous Multi-Pathway Inhibition
Studies reveal that BMP-mediated induction of the totipotent state in mouse embryonic stem cells (ESCs) is constrained by its cross-activation of FGF, Nodal, and Wnt pathways. When BMP signaling is activated, it initiates these parallel pathways that subsequently compensate and limit BMP's primary effects. Rational inhibition of these cross-activated pathways significantly enhances the efficiency of BMP-mediated cellular reprogramming, increasing the proportion of totipotent cells from 30% to over 80% in certain contexts [15]. This approach demonstrates that targeted multi-pathway inhibition rather than single-pathway manipulation is required to achieve specific cell fate outcomes.
Exploiting Temporal Windows of Competence
The timing of pathway manipulation is as critical as the manipulation itself. Research shows that FGF and Nodal pathways mediate cardiac specification during a brief developmental window of just 1-2 hours during early gastrulation, while sustained ERK activation (downstream of FGF) is required for at least four hours to commit cells to the cardiac lineage [56]. Similarly, the BMP/Nodal ratio during specific temporal windows determines whether neuroectoderm or mesoderm drives convergent extension movements [22]. These findings highlight the importance of precisely timed interventions that account for the dynamic nature of signaling interactions rather than continuous pathway manipulation.
Experimental Workflow for Addressing Pathway Redundancy
The following diagram outlines a systematic experimental approach for investigating and overcoming pathway redundancy in gastrulation models:
Figure 2: Experimental workflow for addressing pathway redundancy. This iterative approach combines combinatorial pathway inhibition with temporal precision to achieve specific developmental outcomes.
The Scientist's Toolkit: Essential Research Reagents
Successful investigation of pathway redundancy requires carefully selected experimental tools. The following table summarizes key reagents and their applications in BMP, FGF, and Nodal pathway research:
Table 4: Essential research reagents for studying pathway redundancy in gastrulation
| Reagent Category | Specific Examples | Function/Application | Experimental Context |
|---|---|---|---|
| Small Molecule Inhibitors | SU5402 (FGF receptor inhibitor); SB-431542 (Nodal/Activin inhibitor); XAV939 (Wnt inhibitor); DMH1 (BMP inhibitor) | Targeted pathway inhibition; Testing compensation mechanisms | EpiSC differentiation; Gastruloid models; Zebrafish explants [9] [15] [57] |
| Recombinant Ligands | BMP4 (proximal mesoderm); FGF2/FGF4 (distal mesoderm); Nodal/Activin A (mesendoderm) | Direct pathway activation; Dose-response studies | Concentration-dependent fate mapping; EpiSC differentiation protocols [9] [58] |
| Genetic Tools | Dominant-negative receptors (ΔFGFR1, ΔActRI); Conditional knockout models; Inducible expression systems | Cell-autonomous pathway blockade; Temporal control | Mouse transgenic models; ES cell differentiation; Lineage tracing [56] [55] |
| Reporting Systems | T/Bra:mCherry mesoderm reporter; Phospho-Smad1/5/8 antibodies; ERK/MAPK activity reporters | Real-time signaling visualization; Pathway activity quantification | Live imaging of gastruloids; Immunofluorescence; Single-cell RNA sequencing [9] [59] |
| Experimental Models | Mouse gastruloids; EpiSC; Zebrafish explants; Xenopus animal caps | Reduced complexity systems; High-throughput screening | Pathway interaction studies; Temporal manipulation [3] [9] [22] |
Detailed Experimental Protocols
Combinatorial Inhibition in EpiSC Differentiation
This protocol defines a systematic approach for identifying and overcoming compensatory mechanisms during mesoderm differentiation of mouse Epiblast Stem Cells (EpiSCs).
Materials:
- EpiSCs maintained in FAX medium (ActivinA, FGF2, XAV939)
- N2B27 basal differentiation medium
- Small molecule inhibitors: DMH1 (BMP inhibitor, 1 µM), SU5402 (FGF inhibitor, 50 µM), SB-431542 (Nodal inhibitor, 75 µM), XAV939 (Wnt inhibitor, 2 µM)
- Recombinant proteins: BMP4 (0-100 ng/mL), FGF2 (0-100 ng/mL)
- Fixation buffer: 4% PFA in PBS
- Antibodies: anti-T/BRA (pan-mesoderm), anti-HAND1 (proximal mesoderm), anti-TBX6 (distal mesoderm)
Procedure:
- Maintenance Culture: Maintain EpiSCs in FAX medium on fibronectin-coated plates until 80% confluent.
- Initiate Differentiation: Wash cells twice with N2B27 medium and switch to differentiation medium containing 1 µM Chir99021 (Wnt agonist) and 8 ng/mL BMP4.
- Experimental Conditions:
- Condition A: BMP4 only (8 ng/mL)
- Condition B: BMP4 + SU5402 (50 µM)
- Condition C: BMP4 + SB-431542 (75 µM)
- Condition D: BMP4 + SU5402 + SB-431542 + XAV939
- Condition E: FGF2 only (50 ng/mL)
- Condition F: FGF2 + DMH1 (1 µM)
- Temporal Manipulation: For timing experiments, add inhibitors at different time points (0h, 12h, 24h, 36h post-differentiation) to identify critical windows.
- Harvest and Analysis:
- At 72h post-differentiation, fix cells for immunocytochemistry (T/BRA, HAND1, TBX6).
- Harvest RNA for qPCR analysis of marker genes (Hand1, Gata6, Tbx6, Msgn1, Foxa2).
- For single-cell RNA sequencing, dissociate cells and process using 10X Genomics platform.
Interpretation: Compare expression patterns across conditions to identify compensatory mechanisms. For example, if BMP inhibition alone does not block proximal mesoderm formation, but BMP+FGF inhibition does, this indicates FGF compensation. Similarly, persistence of distal markers despite FGF inhibition suggests BMP or Nodal compensation [9] [15].
Temporal Manipulation of BMP/Nodal Ratios in Gastruloids
This protocol describes how to manipulate the timing of BMP and Nodal pathway activation in mouse gastruloids to control tissue-specific morphogenesis.
Materials:
- Mouse embryonic stem cells (mESCs)
- Gastruloid formation medium: N2B27 with B27 supplement without Vitamin A
- Recombinant proteins: BMP4 (1-100 ng/mL), Nodal/Activin A (10-100 ng/mL)
- Small molecule inhibitors: LDN-193189 (BMP inhibitor, 100 nM), SB-431542 (Nodal inhibitor, 75 µM)
- Agarose-coated 96-well U-bottom plates for gastruloid culture
- Live imaging setup with temperature and CO₂ control
Procedure:
- Gastruloid Formation:
- Harvest mESCs and prepare single-cell suspension at 3×10⁵ cells/mL in gastruloid medium.
- Plate 300 cells (1 µL) in each well of agarose-coated 96-well U-bottom plates.
- Centrifuge plates at 300×g for 3 min to aggregate cells.
- Culture for 96h, changing medium at 48h.
Temporal Signaling Manipulation:
- Group 1: Add BMP4 (20 ng/mL) at 0h, add Nodal (50 ng/mL) at 24h (High→Low BMP/Nodal)
- Group 2: Add Nodal (50 ng/mL) at 0h, add BMP4 (20 ng/mL) at 24h (Low→High BMP/Nodal)
- Group 3: Simultaneous addition of BMP4 (20 ng/mL) and Nodal (50 ng/mL) at 0h
- Group 4: Control with no additions
Combinatorial Inhibition:
- Group 5: BMP4 (20 ng/mL) at 0h + LDN-193189 (100 nM) at 12h
- Group 6: Nodal (50 ng/mL) at 0h + SB-431542 (75 µM) at 12h
Live Imaging and Analysis:
- Image gastruloids every 2h for 72h using brightfield microscopy.
- Quantify elongation ratio (length/width), symmetry breaking, and bud formation.
- Fix at endpoint for immunostaining of tissue-specific markers.
Interpretation: Compare elongation patterns and marker expression across timing groups. Gastruloids with early high BMP/Nodal ratio typically show enhanced neuroectoderm extension, while early low BMP/Nodal ratio promotes mesoderm-driven elongation. Inhibition studies reveal the timing of competence windows for each pathway [3] [22].
Overcoming pathway redundancy in mouse gastrulation research requires a paradigm shift from single-pathway manipulation to network-level interventions. The strategies outlined in this technical guide—combinatorial pathway inhibition, temporal targeting of competence windows, and exploitation of natural antagonisms—provide a framework for achieving specific developmental outcomes despite robust compensatory mechanisms. The experimental protocols and reagents detailed here offer researchers validated approaches to dissect and manipulate these complex signaling networks. As the field advances, leveraging computational modeling informed by quantitative single-cell data and sophisticated in vitro models like gastruloids will be essential for predicting and testing interventions that account for the inherent redundancy of developmental signaling systems [3] [9]. Implementation of these approaches will not only advance basic research but also inform therapeutic strategies for regenerative medicine and developmental disorders where precise control of cell fate decisions is paramount.
Optimizing Small-Molecule Inhibition Cocktails for Specific Pathway Targeting
The process of mouse gastrulation is a precisely orchestrated developmental event governed by the dynamic interplay of key signaling pathways, including Bone Morphogenetic Protein (BMP), Nodal (a TGF-β superfamily member), and Fibroblast Growth Factor (FGF). These pathways form an intricate signaling network that directs the emergence of the three germ layers—ectoderm, mesoderm, and endoderm—and establishes the primary body plan. In this context, small-molecule inhibitors have emerged as indispensable tools for dissecting this complex web of interactions. Unlike genetic knockouts, which are often lethal at these early stages, small molecules offer temporal and dose-dependent control, allowing researchers to inhibit specific pathways at precise developmental windows. The strategic combination of these inhibitors into targeted cocktails enables the systematic interrogation of pathway functions and their contributions to cell fate specification. This guide provides a detailed technical framework for developing and applying these inhibition strategies within the specific experimental context of mouse gastrulation, offering optimized protocols, quantitative data, and visualization tools to advance research in this field.
Core Signaling Pathways and Their Inhibition
During mouse gastrulation, BMP, Nodal/TGF-β, and FGF signaling perform critical, non-overlapping functions. BMP signaling is pivotal for the specification of ventral-posterior cell fates and the establishment of the primordial germ cell lineage. Its activity is balanced by antagonists in the anterior region, which protect the neuroectoderm from BMP-mediated repression. The Nodal/Activin/TGF-β pathway is a master regulator of the primitive streak, mesendoderm formation, and left-right asymmetry. FGF signaling, conversely, is primarily involved in epithelial-to-mesenchymal transition (EMT), a process essential for the ingression of cells through the primitive streak and their subsequent migration to form the mesoderm layer.
The table below summarizes the core components, functions, and specific inhibitors for each of these critical pathways.
Table 1: Core Signaling Pathways in Mouse Gastrulation and Their Inhibitors
| Pathway | Core Ligands & Receptors | Primary Intracellular Effectors | Key Roles in Gastrulation | Common Small-Molecule Inhibitors |
|---|---|---|---|---|
| BMP | BMP2, BMP4; BMPR1A/B (ALK3/6), BMPR2 | SMAD1/5/8, SMAD4 | Ventral-posterior patterning, extraembryonic lineage commitment [60] | LDN193189, Dorsomorphin |
| Nodal/TGF-β | Nodal, Activin; TGFβR1 (ALK5), ACVR1B (ALK4), ACVR1C (ALK7) | SMAD2/3, SMAD4 | Primitive streak formation, mesendoderm induction, left-right asymmetry [60] | SB431542, A83-01 |
| FGF | FGF4, FGF8; FGFR1, FGFR2 | RAS/MAPK, PI3K/AKT | Epithelial-to-mesenchymal transition (EMT), mesoderm migration and specification | PD173074, SU5402, LY294002 |
Mechanism of Dual SMAD Inhibition
The "Dual SMAD Inhibition" strategy, a cornerstone of modern stem cell and developmental biology, provides a powerful illustration of cocktail-based pathway targeting. This approach involves the simultaneous inhibition of both the TGF-β/Activin/Nodal (SMAD2/3) and BMP (SMAD1/5/8) signaling branches [60]. Developmentally, active TGF-β and BMP signaling maintains pluripotency or diverts cells toward mesodermal and endodermal fates while actively repressing neural identity within the ectoderm. By blocking these signals, cells effectively default to a neuroectodermal lineage. This is achieved mechanistically by using small molecules that target specific receptor kinases:
- SB431542: A cell-permeable molecule that selectively inhibits ALK4, ALK5, and ALK7, thereby suppressing the phosphorylation and activation of SMAD2/3 and downstream TGF-β/Nodal signaling events [60].
- LDN193189 / Dorsomorphin: These inhibitors target the intracellular BMP pathway by inhibiting ALK2, ALK3, and ALK6 receptors, which blocks the phosphorylation of SMAD1/5/8 and subsequent transcription of BMP-target genes [60].
The synergistic effect of this cocktail results in highly efficient and reproducible induction of neural fate from pluripotent cells, demonstrating the power of combined pathway inhibition for directing specific developmental outcomes.
Visualizing the Signaling Network and Inhibition Strategy
The following diagram illustrates the core signaling pathways targeted during mouse gastrulation and the points of inhibition for key small molecules.
Quantitative Profiling of Small-Molecule Inhibitors
Selecting the optimal inhibitors requires a careful balance of potency, selectivity, and cost. The following table provides a quantitative comparison of commonly used small molecules for targeting the BMP, Nodal/TGF-β, and FGF pathways in the context of mouse embryogenesis and stem cell differentiation studies.
Table 2: Quantitative Profile of Key Small-Molecule Inhibitors
| Inhibitor | Primary Target(s) (IC₅₀) | Common Working Concentration (in vitro) | Solubility | Key Off-Target Effects | Typical Application in Gastrulation Studies |
|---|---|---|---|---|---|
| LDN193189 | ALK2 (0.6 nM), ALK3 (1.2 nM) [60] | 0.1 - 1 µM | DMSO | Moderate selectivity over ALK5 | Inhibits BMP-SMAD to promote neural/default fate; study ventral patterning. |
| Dorsomorphin | ALK2 (46 nM), ALK3 (108 nM), AMPK | 1 - 10 µM | DMSO | Potently inhibits AMPK | Early-generation BMP inhibitor; used in ventral fate repression studies. |
| SB431542 | ALK4 (94 nM), ALK5 (47 nM), ALK7 (< | 5 - 10 µM | DMSO | Highly selective over ALK1/2/3/6 | Inhibits Nodal/TGF-β-SMAD; essential for mesendoderm induction studies. |
| A83-01 | ALK4 (45 nM), ALK5 (12 nM), ALK7 (7.5 nM) | 0.5 - 5 µM | DMSO | Potent against all TGF-β type I receptors | Alternative to SB431542; more potent pan-TGF-β inhibitor. |
| PD173074 | FGFR1 (25 nM), FGFR2 (100 nM) | 50 - 500 nM | DMSO | Selective for FGFR over other RTKs | Studies on EMT, mesoderm migration, and primitive streak ingression. |
| SU5402 | FGFR1 (20 nM), VEGFR (10 µM) | 5 - 20 µM | DMSO | Inhibits VEGFR at higher concentrations | Functional analysis of FGF signaling in cell fate specification. |
| LY294002 | PI3K (1.4 µM) | 5 - 20 µM | DMSO | Does not distinguish between PI3K isoforms | Targets FGF-PI3K/AKT axis; studies on cell survival/proliferation. |
Experimental Design and Workflow Optimization
A Generalized Experimental Workflow
A systematic approach is required to design and test a small-molecule inhibition cocktail for a specific hypothesis related to mouse gastrulation signaling. The following diagram outlines a robust, iterative workflow for this process.
Detailed Methodologies for Key Assays
4.2.1 Quantitative RT-PCR (qPCR) for Lineage Markers
This protocol is critical for assessing the molecular efficacy of inhibition cocktails by quantifying changes in the expression of key lineage-specific markers.
- RNA Extraction: Culture cells or process embryonic tissue in TRIzol reagent. Use 1 ml TRIzol per 5-10 x 10⁶ cells. Homogenize thoroughly, incubate for 5 minutes, then add 0.2 ml chloroform per 1 ml TRIzol. Shake vigorously, incubate for 3 minutes, and centrifuge at 12,000 x g for 15 minutes at 4°C. Transfer the colorless upper aqueous phase to a new tube [61].
- cDNA Synthesis: Precipitate RNA with 0.5 ml isopropanol per 1 ml TRIzol. Wash the pellet with 75% ethanol and resuspend in RNase-free water. Quantify RNA concentration using a spectrophotometer (e.g., NanoDrop). Reverse transcribe 2 μg of total RNA into cDNA using a kit such as ReverTra Ace with oligo-dT primers [61].
- qPCR Reaction: Perform quantitative PCR using SYBR Green Premix on a real-time PCR machine (e.g., Bio-Rad CFX96). Use a standard 20 μl reaction volume with the following cycling conditions: 95°C for 3 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. Include a melt curve analysis to confirm amplification specificity. Primer Sequences (Mouse):
- Brachyury (T): F-5'-AGCTACAACCAGCAGCACTC-3', R-5'-GGTCTGGTGAGTGGCAATCT-3'
- Sox1: F-5'-CGACGCACATTCACCAGAC-3', R-5'-CGGTACTTGTAATCCGGTGTG-3'
- FoxA2: F-5'-CGGACAAGGAACCAGCTTAC-3', R-5'-TCTTCATGCCGTCCTTCTTC-3'
- GAPDH: F-5'-AGGTCGGTGTGAACGGATTTG-3', R-5'-TGTAGACCATGTAGTTGAGGTCA-3' (Used for normalization) [61].
4.2.2 Immunohistochemistry (IHC) for Protein Localization
This protocol visualizes the expression and subcellular localization of key proteins (e.g., phosphorylated SMADs) in response to inhibition.
- Fixation and Permeabilization: Wash cells cultured on glass coverslips or embryonic tissue sections with PBS. Fix with 4% Paraformaldehyde (PFA) for 30 minutes at room temperature. Wash again with PBS, then permeabilize and block with PBS containing 0.1% Triton X-100 and 5% normal goat/donkey serum for 1 hour at room temperature [61].
- Antibody Staining: Incubate samples with primary antibody diluted in blocking solution overnight at 4°C. Example Antibodies and Dilutions:
- Anti-phospho-SMAD1/5/8 (1:500)
- Anti-SOX2 (1:400)
- Anti-FOXA2 (1:300)
- Anti-Brachyury (T) (1:250)
- The following day, wash samples 3 times with PBS for 5 minutes each. Incubate with compatible Alexa Fluor-conjugated secondary antibodies (e.g., Alexa Fluor 488, 555, or 647) diluted 1:1000 in blocking solution for 1 hour at room temperature, protected from light.
- Nuclear Counterstaining and Mounting: Wash samples 3 times with PBS. Perform nuclear counterstaining with DAPI (5 μg/mL) for 5 minutes. Perform a final PBS wash and mount coverslips onto glass slides using an anti-fade mounting medium. Image using a fluorescence or confocal microscope [61].
The Scientist's Toolkit: Essential Research Reagents
A successful inhibition study relies on a core set of high-quality reagents and materials. The following table details essential components for setting up these experiments.
Table 3: Essential Research Reagent Solutions
| Reagent/Material | Specifications & Recommended Examples | Primary Function in Experiment |
|---|---|---|
| Small-Molecule Inhibitors | >98% purity, validated activity (e.g., from Selleck Chemicals, Tocris, STEMCELL Technologies) | Selective chemical inhibition of specific kinase targets (e.g., ALKs, FGFRs). |
| Cell Culture Medium | Chemically defined, serum-free (e.g., N2B27, mTeSR1 for PSCs) [61] | Provides a defined, reproducible base medium for differentiation and inhibition studies. |
| Extracellular Matrix | Growth Factor Reduced Matrigel, Geltrex, Laminin-521 | Coats culture surfaces to support the attachment and survival of sensitive pluripotent or differentiating cells. |
| Fixative | 4% Paraformaldehyde (PFA) in PBS [61] | Preserves cellular morphology and protein epitopes for subsequent immunohistochemistry analysis. |
| Permeabilization Buffer | PBS with 0.1-0.3% Triton X-100 [61] | Permeabilizes cell membranes to allow antibodies to access intracellular targets. |
| Blocking Solution | PBS with 5% normal serum (e.g., goat, donkey) and 0.1% Triton X-100 | Blocks non-specific antibody binding to reduce background signal in IHC. |
| RNA Extraction Kit | TRIzol reagent or column-based kits (e.g., RNeasy) [61] | Isolates high-quality, intact total RNA for downstream gene expression analysis. |
| cDNA Synthesis Kit | ReverTra Ace or equivalent reverse transcriptase kits [61] | Synthesizes complementary DNA (cDNA) from an RNA template for qPCR. |
| qPCR Master Mix | SYBR Green Premix (e.g., from Bio-Rad, Thermo Fisher) [61] | Contains enzymes, dNTPs, and fluorescent dye for sensitive and quantitative PCR detection. |
| Validated Antibodies | Phospho-specific SMADs, lineage transcription factors (e.g., from Cell Signaling Technology, R&D Systems) | Detects and visualizes specific protein targets and their activation states via IHC/Western blot. |
Addressing Species-Specific Differences in Signaling Responses
Within the context of a broader thesis on signaling pathways in mouse gastrulation, this technical guide addresses a fundamental challenge in biomedical research: the critical species-specific differences in signaling responses between mouse models and humans. Researchers and drug development professionals routinely utilize mouse models to study conserved signaling pathways—including Bone Morphogenetic Protein (BMP), Nodal, and Fibroblast Growth Factor (FGF)—that orchestrate mammalian gastrulation. Although humans and mice share approximately 90% of their genome in regions of conserved synteny and a high degree of amino acid identity in orthologous proteins, signaling responses frequently diverge due to differences in gene expression networks, pathway regulation, and cellular context [62] [63]. These divergences contribute to the high attrition rate of compounds in clinical trials, underscoring the necessity to critically evaluate the translational relevance of mouse models [62]. This guide provides a detailed framework for identifying, quantifying, and accounting for these differences through structured data comparison, specialized experimental protocols, and validated reagent toolkits, thereby enhancing the predictive value of preclinical research.
Core Signaling Pathways in Mouse Gastrulation: A Comparative Baseline
During mouse gastrulation, secreted signaling molecules from the Bmp, Wnt, Nodal, and Fgf families induce and pattern the primitive streak, initiating the cellular rearrangements that generate the definitive body plan [3]. The precise spatiotemporal dynamics of these pathways direct lineage specification and morphogenesis. Understanding the core functions of these pathways in the mouse provides the essential baseline for cross-species comparison.
BMP Signaling: The BMP pathway is integral for the specification of primordial germ cells (PGCs) and the formation of extraembryonic mesoderm. In mouse PGCs, quantitative analysis at single-cell resolution has revealed a characteristically lower BMP response (measured by nuclear pSMAD1/5/9) compared to the surrounding somatic cells in the allantois [64]. Mutations in genes encoding
Bmp4,Bmp8, andBmp2, or their downstream effectorsSmad1andSmad5, lead to a severe reduction or complete loss of PGCs in mice [64].Nodal/Activin Signaling: A component of the broader TGF-β superfamily, Nodal signaling is essential for primitive streak formation and the establishment of laterality. Studies modulating this pathway in preimplantation development show that inhibition with
SB431542can influence the composition of the inner cell mass [65].FGF Signaling: The FGF pathway, acting through the MAPK cascade, is critical for somatic germ layer specification, the epithelial-to-mesenchymal transition (EMT) during gastrulation, and cell migration [64]. Mouse PGCs and surrounding somatic cells exhibit a comparable MAPK response at the initial stages of specification, though this response decreases over time in both cell types [64].
Wnt Signaling: Wnt signaling is required for the specification of both somatic and germ cell fates. In mouse PGC specification, a Wnt response is observed even in the absence of exogenously added ligand, indicating the presence of endogenous Wnt signaling. This response decreases as differentiation proceeds, a dynamic thought to be crucial for PGCs to block a somatic differentiation trajectory [64].
The following table summarizes key quantitative responses of these pathways in mouse models, providing a reference for cross-species analysis.
Table 1: Quantitative Signaling Responses in Mouse Embryonic Development
| Signaling Pathway | Biological Context | Measured Parameter | Response Level / Outcome | Citation |
|---|---|---|---|---|
| BMP | PGC vs. Somatic Cell Specification | Nuclear pSMAD1/5/9 | Significantly lower in SOX2+ AP2γ+ PGCs | [64] |
| FGF/MAPK | PGC Specification | Spry4 reporter activity (Venus) | Reduced over time; initially no significant difference between PGCs and non-PGCs | [64] |
| Wnt | PGC Specification | TCF/Lef:H2B-GFP reporter | Heterogeneous response; decreases during differentiation | [64] |
| Nodal/Activin | Preimplantation Development | Blastocyst development rate | Inhibition (SB431542, 10μM): 25% vs. Control: 28% | [65] |
Quantitative Comparison of Signaling Responses
A systematic, quantitative approach is essential for delineating species-specific signaling differences. Advanced single-cell technologies now enable the precise measurement of pathway activity across species and developmental stages.
Analytical Techniques for Single-Cell Resolution
Mapping signaling responses at single-cell resolution over time is a powerful strategy for uncovering heterogeneity and divergent activities that bulk analyses would obscure.
- Reporter Cell Lines: The generation of mouse embryonic stem cells (mESCs) with endogenous reporters for immediate-early pathway targets allows for real-time tracking. Key examples include
Spry4H2BVenusfor MAPK/ERK signaling andTCF/Lef:H2B-GFPfor Wnt/β-catenin signaling [64]. - Quantitative Immunofluorescence: This method provides precise, protein-level quantification of signaling activity. For instance, the measurement of nuclear localized phosphorylated SMAD1/5/9 (pSMAD1/5/9) enables the direct comparison of BMP pathway response between distinct cell types, such as PGCs and surrounding somatic cells [64].
- Single-Cell RNA Sequencing (scRNA-seq): Large-scale scRNA-seq atlases, such as one profiling 12.4 million nuclei from 83 mouse embryos, provide a foundational reference for transcriptional states and lineage relationships [29]. Comparing these maps with emerging human embryo data can reveal differences in pathway gene expression and co-expression networks.
Table 2: Conserved and Divergent Features of Human and Mouse Biology
| Aspect | Degree of Conservation | Key Findings | Implications for Signaling Studies |
|---|---|---|---|
| Genome | High | ~90% conserved synteny; 40% nucleotide alignment; 78.5% amino acid identity in orthologs [62] [63] | High conservation of core pathway components (e.g., ligands, receptors). |
| Gene Co-expression | Variable by tissue/process | Brain and bone-associated genes are strongly conserved; testis, eye, skin, and PI3K signaling genes are more divergent [63] | Pathway outcomes may differ in divergent tissues/processes despite conserved genes. |
| dN/dS Ratio vs. Co-expression | Negative Correlation | A negative correlation (rho = -0.19) exists between dN/dS and conservation of co-expression connectivity [63] | Genes with high molecular evolution rates are more likely to have divergent regulatory networks. |
| Disease Modeling | Variable efficacy | Co-expression of metabolic disorder genes is highly conserved; tumor-related genes are the most divergent [63] | Mouse models are more predictive for conserved processes like metabolism than for some cancers. |
Key Divergences in Pathway Activity and Function
Comparative analyses have pinpointed specific pathways and tissues where human and mouse signaling responses notably diverge.
- PI3K-AKT-mTOR Pathway: This critical signaling cascade shows significant divergence between humans and mice. Surprisingly, the most crucial nodes of the pathway, including
mTORandAKT2, exhibit differential co-expression connectivity, suggesting fundamental differences in network regulation that could impact studies of growth, metabolism, and cancer [63]. - Hippo Signaling Pathway: While Hippo is a key regulator of trophectoderm (TE) differentiation in both species, studies indicate "notable species-specific differences" in its role during the first lineage specification in preimplantation development [65]. The precise molecular mechanisms governing YAP/TAZ localization and activity can vary.
- Tissue-Specific Divergence: Genes expressed in the testis, eye, and skin, as well as those involved in olfaction and immune responses, display higher divergence in co-expression networks [63]. This suggests that signaling pathways operating in these tissues are more likely to exhibit species-specific behaviors.
Experimental Protocols for Cross-Species Validation
To ensure the fidelity of mouse models, researchers must employ rigorous experimental designs that directly test the conservation of signaling mechanisms. The following protocols provide a framework for such validation.
Protocol: Quantitative Single-Cell Analysis of Signaling Responses In Vitro
This protocol, adapted from a study on mouse PGC specification, details how to measure and compare BMP, MAPK, and WNT signaling activities in vitro [64].
Cell Line Generation:
- Utilize reporter mESC lines, such as
Spry4H2BVenus(MAPK response) andTCF/Lef:H2B-GFP(WNT response). - Differentiate mESCs into the desired cell type (e.g., PGC-like cells, PGCLCs) under defined conditions, typically involving aggregation and specific cytokine cocktails (BMP4, SCF, EGF, and LIF) [64].
- Utilize reporter mESC lines, such as
Stimulation and Fixation:
- At defined time points during differentiation (e.g., days 2, 4, and 6), harvest cell aggregates.
- Fix cells in 4% paraformaldehyde (PFA) for 15-20 minutes at room temperature.
Immunostaining:
- Permeabilize fixed cells with 0.5% Triton X-100 for 20 minutes.
- Block nonspecific sites with 5% bovine serum albumin (BSA) for 1 hour.
- Incubate with primary antibodies overnight at 4°C. Key antibodies include:
- Anti-pSMAD1/5/9 (BMP signaling response)
- Anti-SOX2 and Anti-AP2γ (for identifying PGCLC population)
- Wash and incubate with appropriate fluorescently conjugated secondary antibodies for 1-2 hours at room temperature.
Imaging and Quantification:
- Image stained aggregates using confocal microscopy, capturing z-stacks to ensure complete cellular coverage.
- Use image analysis software (e.g., ImageJ, CellProfiler) to perform single-cell segmentation and quantify mean fluorescence intensity of the signaling reporter (Venus/GFP) or immunostaining (pSMAD) within individual nuclei.
- Statistically compare signaling responses between different cell populations (e.g., SOX2+ AP2γ+ PGCLCs vs. AP2γ- non-PGCLCs) using a non-parametric test like the Mann-Whitney U test, as implemented in the original study [64].
Protocol: Validating Signaling Requirements via Mutant Chimeras
This protocol, derived from a single-embryo study of mouse gastrulation, describes a powerful in vivo method for testing the function of a signaling component over time and across lineages [24].
Mutant Cell Generation:
Chimera Production:
- Inject a controlled number of the mutant mESCs into wild-type host mouse blastocysts.
- Alternatively, use tetraploid complementation assays, where mutant mESCs are aggregated with tetraploid wild-type embryos, resulting in fetuses derived almost entirely from the mutant ES cells.
Single-Embryo scRNA-seq and Analysis:
- Collect and precisely stage individual chimeric embryos at multiple time points based on somite count or morphological criteria [29] [24].
- Process each embryo individually for scRNA-seq using a high-throughput method like single-cell combinatorial indexing (sci-RNA-seq3) [29].
- Analyze the resulting data to infer "differentiation flows and lineage specification dynamics" [24]. Specifically, assess the contribution and transcriptional state of mutant cells compared to wild-type cells across different lineages and time points, thereby determining the cell-autonomous and non-autonomous requirements for the signaling component.
Figure 1: A unified experimental workflow for addressing species-specific signaling differences, integrating in vitro and in vivo validation strategies.
The Scientist's Toolkit: Research Reagent Solutions
The following table catalogs essential reagents and tools, derived from the cited research, for conducting experiments on signaling pathways in mouse models and for cross-species comparison.
Table 3: Key Research Reagents for Signaling Pathway Analysis
| Reagent / Tool | Function / Target | Application in Context | Example Usage |
|---|---|---|---|
| CRT0276121 | Activator of Hippo signaling | Studies of trophectoderm (TE) vs. inner cell mass (ICM) lineage specification in preimplantation embryos [65] | Treatment from pre-compaction to blastocyst stage at 1.5μM [65] |
| PD0325901 | Inhibitor of FGF/MAPK signaling (MEK inhibitor) | Investigation of FGF role in germ layer specification and PGC formation [65] [64] | Used at 0.5-1.0μM in PGCLC differentiation medium [65] |
| SB431542 | Inhibitor of TGF-β/Activin/Nodal signaling | Analysis of Nodal pathway function in epiblast maturation and lineage priming [65] | Treatment from day 3 to day 6 of in vitro culture at 10μM [65] |
| BMP4 ( recombinant protein) | Activator of BMP signaling (ligand) | Specification of primordial germ cells (PGCs) and extraembryonic mesoderm [64] | Added at 100-500 ng/mL to differentiation media to induce PGC fate [64] |
| Spry4H2BVenus mESCs | Live-cell reporter for MAPK/ERK signaling | Real-time tracking of MAPK activity during cell fate decisions [64] | Quantification of Venus fluorescence in PGCLCs vs. non-PGCLCs [64] |
| TCF/Lef:H2B-GFP mESCs | Live-cell reporter for Wnt/β-catenin signaling | Monitoring endogenous Wnt pathway activity during differentiation [64] | Identification of Wnt-responsive cells in PGCLC aggregates [64] |
| Anti-pSMAD1/5/9 | Antibody for BMP pathway readout | Quantitative immunofluorescence of canonical BMP-SMAD signaling response [64] | Measuring nuclear pSMAD levels to compare BMP activity in PGCs vs. somatic neighbors [64] |
The mouse remains an indispensable model for deconstructing the complex signaling networks that govern gastrulation and early development. However, a critical and nuanced understanding of its limitations is paramount for translational research. By adopting the quantitative, single-cell resolution approaches outlined in this guide—including direct signaling measurement, careful in vitro and in vivo validation, and the use of specialized reagent toolkits—researchers can systematically identify and account for species-specific differences in BMP, Nodal, FGF, and other key pathway responses. This rigorous framework will not only refine the utility of mouse models but also strengthen the foundation upon which our understanding of human development and disease is built, ultimately improving the success rate of translating basic research into clinical applications.
Resolving Technical Noise in Low-Input Epigenomic and Transcriptomic Data
The study of signaling pathways such as BMP, Nodal, and FGF during mouse gastrulation has been profoundly advanced by single-cell technologies. These methods enable genome- and epigenome-wide profiling of thousands of individual cells, offering unprecedented resolution to uncover the molecular events that guide the formation of the body plan [66] [3]. However, the frequent necessity of working with low-input cellular material—a common scenario in embryonic research—exacerbates the challenge of technical noise, including dropout events (where transcripts fail to be detected) and batch effects [66] [65]. These artifacts obscure subtle biological signals, such as the initial fluctuations in pathway activity that precede germ layer specification, hindering the detection of rare cell types and compromising cross-dataset comparisons [66]. This technical guide outlines robust statistical and computational strategies to mitigate these challenges, ensuring that high-resolution biological structures, particularly those involved in key gastrulation signaling networks, can be revealed with greater fidelity.
Understanding Technical Noise and Its Impact on Signaling Studies
Technical noise in single-cell data arises from the entire process of data generation, from cell lysis to sequencing. This is characterized by non-biological fluctuations in molecule detection rates, leading to sparse data matrices where true expression signals are missing or obscured [66]. In low-input protocols, such as those used for rare embryonic cell populations, this problem is intensified due to the limited starting material.
- The Curse of Dimensionality: Single-cell data are inherently high-dimensional, and the accumulation of technical noise across thousands of genes and genomic loci distorts the underlying data structure. It has been mathematically demonstrated that standard dimensionality reduction techniques like PCA are insufficient to overcome this problem [66].
- Impact on Signaling Pathway Analysis: Technical noise can mask critical biological phenomena. In developmental contexts, high dropout rates have been shown to obscure cell-type-specific transcription factor activities and subtle expression changes in key pathway genes, such as Bmp4, Nodal, or Fgf8, which are crucial for patterning the primitive streak [66] [3]. Furthermore, batch effects introduced by processing samples at different times or with different reagents can create non-biological clusters that may be misinterpreted as distinct developmental states [66].
The diagram below illustrates how technical noise and batch effects confound biological signals in single-cell data and the process of noise reduction.
The RECODE Platform: A Comprehensive Solution
RECODE (Resolution of the Curse of Dimensionality) is a high-dimensional statistics-based tool specifically designed to address technical noise and batch effects while preserving the full dimensionality of single-cell data. Its recent upgrade, iRECODE, enables the simultaneous reduction of both technical and batch noise, making it particularly suited for integrative studies across multiple experiments or conditions [66] [67].
Core Algorithm and Workflow
The RECODE algorithm does not rely on imputation. Instead, it models technical noise from the entire data generation process as a general probability distribution and reduces it using eigenvalue modification theory rooted in high-dimensional statistics [66]. The workflow involves three key steps:
- Noise Variance-Stabilizing Normalization (NVSN): The raw gene expression data is mapped to an essential space using NVSN, which stabilizes the variance of technical noise across the dynamic range of expression [66].
- Singular Value Decomposition (SVD): The normalized data is decomposed via SVD to separate signal from noise in a lower-dimensional essential space [66].
- Principal Component Variance Modification and Elimination: The variances of the principal components are modified to suppress components dominated by technical noise, thereby recovering the underlying biological signal [66].
For batch correction, iRECODE integrates established methods like Harmony, MNN-correct, or Scanorama within this essential space. This strategy avoids the high-dimensional calculations that degrade the accuracy and efficiency of most batch-correction methods, enabling simultaneous noise reduction with low computational cost [66].
Table 1: Performance Metrics of iRECODE in scRNA-seq Denoising and Batch Integration
| Metric | Raw Data | RECODE Only | iRECODE (with Harmony) |
|---|---|---|---|
| Relative Error in Mean Expression | 11.1% - 14.3% | Not Available | 2.4% - 2.5% |
| Genomic Scale Relative Error | Baseline (Highest) | Improved vs. Raw | >20% Improvement vs. Raw; >10% Improvement vs. RECODE |
| Batch Mixing (iLISI Score) | Low | Not Available | High, comparable to Harmony |
| Cell-type Identity (cLISI Score) | High | Not Available | Preserved, stable |
| Computational Efficiency | N/A | N/A | ~10x faster than sequential noise reduction + batch correction |
Protocol: Applying iRECODE to scRNA-seq Data
This protocol is adapted from the methodology detailed by Imoto [66].
Input Data Preparation:
- Format your raw count matrix (cells x genes) and metadata specifying batch and experimental conditions.
- Ensure data is from a consistent technology (e.g., all 10x Genomics or all Smart-seq2) for optimal results.
Noise Variance-Stabilizing Normalization (NVSN):
- Apply NVSN to the raw count matrix. This step models the technical noise as a negative binomial distribution and stabilizes its variance across different expression levels.
- Code (R/Python): Utilize the RECODE package function for
NVSN_transform().
Singular Value Decomposition (SVD):
- Perform SVD on the NVSN-transformed matrix to obtain the essential space representation.
- Code:
svd_result <- svd(nvsn_matrix)
Principal Component Variance Modification:
- Apply the RECODE core algorithm to modify the variances of the principal components from the SVD, eliminating components dominated by technical noise.
- Code: Use the
RECODE_denoise()function on thesvd_result.
Integrative Batch Correction with iRECODE:
- Within the denoised essential space, apply the integrated batch correction method (e.g., Harmony). The function will minimize batch effects while preserving biological variation.
- Code: Call
iRECODE_integrate()specifying the batch covariate and choosing Harmony as the backend.
Output and Downstream Analysis:
- The output is a full-dimensional, denoised, and batch-corrected count matrix.
- This matrix can be used directly for downstream analyses like clustering, trajectory inference, and differential expression of key gastrulation genes (e.g., Brachyury, Eomes, Foxa2).
Application to Epigenomic and Spatial Transcriptomic Data
The principles behind RECODE make it applicable beyond scRNA-seq to any single-cell data generated by random molecular sampling, including epigenomic assays like scATAC-seq and scHi-C, as well as spatial transcriptomics [66].
Denoising Single-Cell Hi-C Data
scHi-C data, which captures chromosome conformation, is notoriously sparse. RECODE can be applied to the vectorized upper triangle of scHi-C contact maps [66].
- Workflow: The NVSN distribution confirms that scHi-C data affected by technical noise is amenable to RECODE processing. Application of RECODE significantly reduces sparsity, aligning scHi-C-derived topologically associating domains (TADs) with their counterparts from bulk Hi-C data, which are considered a gold standard [66].
- Significance for Development: Cleaner scHi-C data enables the more reliable detection of cell-type-specific chromatin interactions that may regulate the expression of key signaling genes during gastrulation.
Enhancing Spatial Transcriptomics
Spatial transcriptomics technologies often have limited resolution, with each capture spot containing transcripts from multiple cells. This requires deconvolution to infer cell-type composition [68]. RECODE can denoise the underlying gene expression matrix before deconvolution, improving the accuracy of subsequent steps.
- Integration with Deconvolution Algorithms: Denoised data from RECODE serves as a superior input for deconvolution tools like cell2location, SPOTlight, or STRIDE [68]. These tools use probabilistic models, non-negative matrix factorization (NMF), or deep learning to infer the proportion of cell types within each spot.
- Protocol for Spatial Data:
- Apply RECODE to the raw spot-by-gene count matrix from platforms like 10x Visium.
- Use the denoised matrix as input to a deconvolution algorithm alongside a single-cell RNA-seq reference atlas.
- The result is a high-resolution map of cell types and states, positioned within the spatial context of the embryo, allowing researchers to visualize how BMP, Nodal, and FGF signaling activities are localized.
Table 2: Research Reagent Solutions for Signaling and Noise Reduction Studies
| Reagent / Tool | Function | Application in Gastrulation Research |
|---|---|---|
| PD0325901 | Inhibitor of FGF signaling (MEK inhibitor) | Used to dissect FGF role in primitive streak patterning [65] |
| SB431542 | Inhibitor of Nodal/TGF-β signaling (ALK4/5/7 inhibitor) | Applied to study Nodal's function in germ layer specification [65] |
| BMP4 (Recombinant) | Activator of BMP signaling | Used in vitro to mimic BMP gradient effects on embryonic patterning [65] |
| TRULI | Inhibitor of the Hippo signaling pathway | Manipulates Hippo pathway to study trophectoderm vs. inner cell mass fate [65] |
| RECODE Software | Algorithm for technical noise and batch effect reduction | Cleans scRNA-seq, scHi-C, and spatial data from low-input embryonic samples [66] |
| cell2location / SPOTlight | Spatial transcriptomics deconvolution algorithms | Infers cell-type composition within spatial spots of embryonic tissue sections [68] |
Integrating Denoised Data into Gastrulation Signaling Research
With technical noise mitigated, denoised data provides a more reliable foundation for modeling the gene regulatory networks that control gastrulation. The core signaling pathways—BMP, Wnt, Nodal, and FGF—interact in a complex spatiotemporal sequence to induce and pattern the primitive streak [3] [65]. The following diagram synthesizes the relationships between these pathways and the key structures of the gastrulating embryo, based on information from mouse studies and gastruloid models [3] [65].
Experimental Framework for Validating Signaling Dynamics:
- Targeted Perturbation: Treat gastruloids or embryonic stem cells with small molecule inhibitors (e.g., SB431542 for Nodal) or recombinant proteins (e.g., BMP4) as listed in Table 2 [65].
- Multi-modal Profiling: Collect scRNA-seq and/or scHi-C data from perturbed and control samples. Low-input protocols may be necessary.
- Data Denoising and Integration: Process all data simultaneously with iRECODE to reduce technical noise and batch effects from the perturbation experiment, creating a clean, integrated dataset.
- Network Inference: Analyze the denoised data to reconstruct the gene regulatory network. Identify key transcription factors downstream of BMP, Nodal, and FGF signaling and their target genes.
- Spatial Validation: Using spatial transcriptomics deconvolution on denoised data, validate the predicted spatial localization of signaling activities and resulting cell fates within the embryonic context [68].
Technical noise is a significant barrier to extracting meaningful biological insights from low-input epigenomic and transcriptomic data. The RECODE platform provides a robust, versatile, and efficient solution for mitigating this noise and batch effects across diverse data modalities. By integrating these computational advances with targeted experimental perturbations in model systems like gastruloids, researchers can achieve an unprecedented, high-resolution view of the signaling dynamics that orchestrate mouse gastrulation. This approach enables a more accurate and integrated understanding of how the BMP, Nodal, FGF, and Wnt pathways interact to generate the mammalian body plan.
Interpreting Complex Phenotypes from Pathway Cross-Activation and Antagonism
The process of mouse gastrulation is orchestrated by a complex interplay of key signaling pathways, primarily BMP, Nodal, and FGF. Recent research has revealed that the cross-activation and antagonistic relationships between these pathways are not merely background noise but fundamental mechanisms that shape cell fate decisions and pattern formation. This technical guide synthesizes current understanding of how these signaling interactions generate complex phenotypes, providing researchers with structured data, experimental protocols, and computational frameworks to advance studies in developmental biology and therapeutic development.
Pathway Interactions: A Network Perspective
Signaling pathways during mouse gastrulation form an intricate network where crosstalk begins at the receptor level and extends through multiple downstream effectors [69]. Growth factor receptors with intrinsic tyrosine kinase activity, for instance, interact with multiple effector pathways including Ras-ERK, PI3K, and phospholipase-Cγ pathways [69]. Similarly, G protein-coupled receptors can transmit signals through both Gα and Gβγ subunits to multiple downstream effectors [69]. This interconnectivity creates a sophisticated information-processing system that enables cells to make fate decisions based on integrated signaling inputs.
Table 1: Core Signaling Pathways in Mouse Gastrulation
| Pathway | Primary Ligands | Main Receptors | Key Transducers | Primary Roles in Gastrulation |
|---|---|---|---|---|
| BMP | BMP4 | BMPR-I/II | SMAD1/5/8, SMAD4 | Proximal mesoderm patterning, totipotent state induction [15] [9] |
| Nodal | Nodal, Activin | Activin receptor | SMAD2/3, SMAD4 | Primitive streak formation, distal mesoderm patterning [9] [3] |
| FGF | FGF4, FGF8 | FGFR1 | FRS2, GRB2, SOS | Distal mesoderm specification, migration control [9] |
| Wnt | Wnt3a | Frizzled, LRP5/6 | β-catenin, TCF/LEF | Posterior patterning, mesoderm induction [15] [3] |
The connectivity within these signaling networks can be represented through various computational frameworks, each offering distinct advantages for understanding information flow. Graph theory provides a mathematical foundation for analyzing these networks, where molecules represent nodes and their interactions form edges [69] [70]. However, traditional graph representations may oversimplify the biological reality of signaling pathways, where reactions often involve multiple inputs and outputs simultaneously [70]. Advanced modeling approaches including hypergraphs, bipartite graphs, and compound graphs offer enhanced capability to capture the many-to-many relationships inherent in biochemical reactions [70].
Quantitative Dynamics of Pathway Interactions
Antagonistic BMP-FGF Relationship
Recent single-cell resolution studies in differentiating epiblast stem cells (EpiSCs) have quantified the opposing functions of BMP and FGF signaling during mesoderm differentiation [9]. This antagonistic relationship follows concentration-dependent principles that can be measured experimentally:
Table 2: BMP4 Concentration-Dependent Fate Patterning in EpiSCs
| BMP4 Concentration (ng/ml) | Primary Marker Expression | Regional Embryonic Equivalent | Cell Fate Outcomes |
|---|---|---|---|
| 0-2 | Foxa2, Shh (peak) | Most distal | Axial mesoderm, definitive endoderm |
| ~4 | T/Bra, Tbx6, Msgn1 (peak) | Distal | Paraxial, presomitic mesoderm |
| 16-32 | Hand1, Gata6 (peak) | Proximal | Extraembryonic, lateral plate mesoderm |
| >64 | Hand1 (sustained) | Most proximal | Extraembryonic mesoderm |
This concentration-dependent patterning establishes BMP as a morphogen in gastrulation, with different threshold concentrations specifying distinct mesodermal subtypes [9]. The antagonism between BMP and FGF manifests clearly in experimental observations that FGF stimulation boosts endogenous Fgf genes while simultaneously repressing Bmp ligand genes [9]. This reciprocal inhibition creates a bistable system that promotes coherent domain specification through a community effect.
Cross-Activation Constraints on BMP Signaling
While BMP signaling alone can induce the totipotent state in mouse embryonic stem cells (ESCs), its efficacy is constrained by cross-activation of other pathways. Research demonstrates that BMP signaling promotes ESC heterogeneity by cross-activating FGF, NODAL, and WNT pathways [15]. This cross-activation creates a balancing mechanism that limits the proportion of cells entering the totipotent state.
Experimental inhibition of these BMP-mediated cross-activations provides compelling evidence for this model. When FGF, NODAL, and WNT pathways are rationally inhibited alongside BMP activation, researchers observe a significant enhancement in totipotent cell proportions [15]. Single-cell mRNA sequencing confirmed that induction of the totipotent state under these conditions was accompanied by suppression of primed and primitive endoderm states [15].
Experimental Models and Methodologies
Stem Cell Differentiation Protocols
EpiSC Differentiation for Mesoderm Patterning Studies
- Starting Culture: Maintain EpiSCs in N2B27 medium supplemented with ActivinA (20ng/ml), FGF2 (12ng/ml), and XAV939 (2µM) to establish a homogeneous, primed pluripotent population [9].
- Mesoderm Induction: Replace ActivinA and XAV939 with Chir99021 (1µM) and varying concentrations of BMP4 (0-100ng/ml) to trigger mesoderm differentiation [9].
- Time Course: Analyze markers at 24-72 hours post-induction using quantitative PCR, immunostaining, or single-cell RNA sequencing.
- Key Readouts: T/BRA (pan-mesoderm), TBX6 (distal mesoderm), HAND1 (proximal mesoderm) protein expression via In-Cell Western or immunofluorescence [9].
Mouse ESC Totipotency Induction Protocol
- Basal Medium: Standard mouse ESC culture conditions.
- Totipotency Induction: Activate BMP signaling (BMP4 at 10-50ng/ml) while simultaneously inhibiting FGF (PD173074 100nM), NODAL (SB431542 10µM), and WNT (IWP2 5µM) pathways [15].
- Validation: Assess totipotency markers (e.g., MERVL, Zscan4) via single-cell RNA sequencing and functional characterization through chimaera formation assays [15].
- Timeline: Peak totipotent cell populations observed at 48-72 hours post-induction.
Signaling History Tracking in Live Cells
Understanding the temporal dynamics of signaling pathways requires sophisticated live-cell imaging approaches:
- Reporter Lines: Generate ESC lines with endogenously tagged signaling components (e.g., GFP::SMAD4, RFP::SMAD1) [10].
- Imaging Setup: Continuous live imaging over 48-72 hours with time-lapse microscopy (30-60 minute intervals) [10].
- Signal Quantification: Calculate nuclear-to-cytoplasmic ratios of fluorescent reporters as a proxy for pathway activity [10].
- Endpoint Analysis: Combine with iterative immunofluorescence staining for fate markers in the same cells after live imaging [10].
- Data Integration: Use computational approaches (e.g., principal component analysis) to classify signaling history clusters and correlate with fate outcomes [10].
Computational Modeling and Visualization
Signaling Network Representations
The complexity of signaling pathway interactions necessitates computational models to understand information processing capabilities. Several network representations offer complementary insights:
Graph-Based Models represent molecules as nodes and interactions as edges, allowing application of graph theory principles to identify network properties [69] [70]. While intuitive, this approach may oversimplify biochemical reactions that inherently involve multiple participants [70].
Hypergraph Models generalize traditional graphs to capture many-to-many relationships, providing more biologically accurate representations of signaling reactions where multiple inputs and outputs occur simultaneously [70]. This approach reveals that Reactome pathways are poorly connected as hypergraphs compared to their graph representations, highlighting the restrictive nature of biochemical connectivity constraints [70].
B-relaxation Distance offers a novel parameterized transition between strict hypergraph connectivity and permissive graph connectivity, enabling researchers to identify subtle structures within pathway topologies and quantify one pathway's downstream influence on another [70].
Visualizing Pathway Interactions
Network of BMP, FGF, Nodal, and WNT pathway crosstalk during gastrulation. Green arrows (BMP) indicate activation, while colored curved arrows show antagonistic relationships.
Experimental Workflow for Signaling Studies
Comprehensive workflow from cell culture through computational modeling for studying signaling pathways.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Signaling Pathway Studies
| Reagent Category | Specific Examples | Function/Application | Concentration Range |
|---|---|---|---|
| Pathway Agonists | BMP4 (ligand), Chir99021 (Wnt agonist) | Activate specific signaling pathways | BMP4: 2-100 ng/ml; Chir99021: 1-3 µM [15] [9] |
| Pathway Antagonists | Dorsomorphin (BMP inhibitor), PD173074 (FGF inhibitor), SB431542 (Nodal inhibitor) | Specifically inhibit target pathways to test functional requirements | Dorsomorphin: 1-10 µM; PD173074: 100-500 nM; SB431542: 10 µM [15] |
| Signaling Reporters | GFP::SMAD4, RFP::SMAD1 knock-in cell lines | Live imaging of pathway activation dynamics | Endogenous tagging for accurate quantification [10] |
| Cell Fate Markers | T/BRA (pan-mesoderm), TBX6 (distal mesoderm), HAND1 (proximal mesoderm) | Identify differentiated cell states via immunostaining or qPCR | Antibody-dependent concentrations [9] |
| Small Molecule Modulators | XAV939 (Wnt inhibitor), SU5402 (FGF inhibitor) | Fine-tune pathway activity levels in concentration-dependent studies | XAV939: 2-5 µM; SU5402: 5-20 µM [9] |
Emerging Concepts and Future Directions
Signaling Dynamics and Temporal Integration
The traditional morphogen gradient model is being supplemented by more sophisticated understandings of signaling dynamics and temporal integration. Research in human pluripotent stem cells demonstrates that BMP signaling history correlates strongly with fate decisions in individual cells [10]. Surprisingly, both the level and duration of BMP signaling activity control cell fate choices primarily by changing the time integral of signaling, suggesting that duration and level are interchangeable in this developmental context [10].
This temporal integration mechanism appears to operate through gradual accumulation or depletion of key regulatory factors. Evidence suggests that SOX2 may serve as a integrator of BMP signaling, with its levels decreasing in proportion to the time integral of BMP activity [10]. This creates a system where lower-level signaling over extended periods can produce equivalent differentiation outcomes to higher-level signaling over shorter durations.
Advanced Model Systems
The field is increasingly leveraging sophisticated in vitro models to study signaling dynamics with enhanced temporal and spatial resolution. Gastruloids - 3D aggregates of embryonic stem cells that recapitulate aspects of gastrulation - provide a powerful platform for observing signaling dynamics in a system that mirrors embryonic patterning [3]. These models, combined with live imaging and computational approaches, offer unprecedented access to the signaling dynamics that shape the embryonic body plan.
Interpreting complex phenotypes arising from pathway cross-activation and antagonism requires integration of multiple conceptual frameworks:
First, pathway interactions follow network principles with emergent properties including ultrasensitivity, bistability, and robust patterning [69]. The demonstrated antagonism between BMP and FGF creates a bistable system that promotes coherent domain specification [9].
Second, signaling dynamics and temporal integration are as critical as concentration gradients in fate determination [10]. The time integral of BMP signaling, rather than instantaneous levels, determines differentiation outcomes.
Third, cross-activation creates constraint mechanisms that balance cell state transitions, as evidenced by BMP-mediated activation of FGF, NODAL, and WNT pathways that ultimately limit the proportion of cells entering the totipotent state [15].
These principles provide researchers with a conceptual toolkit for designing experiments, interpreting complex phenotypes, and developing therapeutic strategies that target developmental signaling pathways in disease contexts. As single-cell technologies and computational modeling approaches continue to advance, our ability to decode the sophisticated language of signaling crosstalk during gastrulation will continue to improve, offering new insights into both normal development and disease processes.
Strategies for Enhancing Signaling-Mediated Cell State Transitions
The precise control of cell state transitions is a fundamental objective in developmental biology and regenerative medicine. This process is governed by a complex interplay of signaling pathways that direct cells toward specific fates. Within the context of mouse gastrulation and embryonic stem cell (ESC) biology, the pathways mediated by Bone Morphogenetic Protein (BMP), NODAL, and Fibroblast Growth Factor (FGF) represent a critical signaling network that determines cellular identity. Recent research has revealed that the cross-activation and mutual antagonism between these pathways create a signaling landscape that can be strategically manipulated to direct cell fate transitions. This technical guide synthesizes current understanding of these interactions and provides detailed methodologies for enhancing signaling-mediated cell state transitions, with particular emphasis on the induction of the totipotent state in mouse ESCs. The insights presented herein are framed within a broader thesis on signaling pathways in mouse gastrulation, offering researchers a comprehensive framework for experimental design and implementation in both developmental biology and therapeutic applications.
Mechanistic Foundations of Pathway Interactions
BMP Signaling as a Primary Inducer of the Totipotent State
BMP signaling has been identified as a potent inducer of the totipotent state in mouse embryonic stem cells (ESCs). Totipotent cells possess the capacity to generate all cell types in an organism, including both embryonic and extra-embryonic tissues. In cultured mouse ESCs, which naturally exist in multiple states resembling distinct stages of early embryogenesis, BMP activation initiates a transcriptional program that drives cells toward this privileged developmental state. However, the efficiency of BMP-mediated induction is constrained by intrinsic regulatory mechanisms that involve cross-activation of other signaling pathways [26] [15].
Single-cell mRNA sequencing analyses have revealed that successful induction of the totipotent state via BMP signaling is accompanied by the concurrent suppression of primed pluripotent and primitive endoderm states. This suggests that BMP-mediated totipotency induction operates through a dual mechanism: both activating totipotency-associated transcriptional programs and actively repressing alternative cell fate trajectories. The resulting reprogrammed totipotent cells generated through BMP activation closely resemble totipotent cells identified in preimplantation mouse embryos at both transcriptional and functional levels [26].
Constraining Mechanisms of Pathway Cross-Activation
The efficacy of BMP signaling in inducing the totipotent state is limited by its tendency to cross-activate three other major signaling pathways: FGF, NODAL, and WNT. This cross-activation creates a self-regulating circuit that maintains ESC heterogeneity by preventing uniform transition to the totipotent state. Research by Thulaj Meharwade and colleagues demonstrated that BMP signaling initiates a cascade that activates FGF, NODAL, and WNT pathways, which in turn constrain the full manifestation of BMP-directed totipotency [26] [15].
This cross-activation network represents a fundamental mechanism regulating stem cell heterogeneity. The balanced co-existence of multiple cellular states within ESC cultures is maintained through this intricate signaling dialogue. From an experimental perspective, this constraint mechanism can be exploited to enhance the proportion of totipotent cells through rational inhibition of the cross-activated pathways, thereby amplifying BMP's totipotency-inducing effects [26].
BMP-NODAL Integration Through Smad Antagonism
The integration of BMP and NODAL signaling occurs through a mechanism of mutual effector antagonism at the level of their downstream Smad transcription factors. In zebrafish embryo studies, it has been demonstrated that Smad2 (the primary transducer of NODAL signaling) and Smad5 (a key transducer of BMP signaling) antagonize each other's functions in a manner that enables cells to precisely interpret overlapping signal gradients [5].
This antagonistic interaction creates a system where cell fate specification is determined not by absolute levels of either signal alone, but by the relative activity ratio of pSmad2 to pSmad5. Cells can thus discriminate between overlapping BMP and NODAL signal gradients and activate distinct transcriptional programs based on this ratio. This mechanistic insight explains how embryonic patterning can emerge from opposing sources of BMP and NODAL signaling molecules, and provides a strategic foundation for manipulating cell fate decisions through controlled modulation of this signaling balance [5].
Table 1: Signaling Pathway Interactions and Functional Outcomes
| Signaling Pathway | Primary Transducers | Cross-Activation Targets | Cell State Influence |
|---|---|---|---|
| BMP | Smad1/5/8 | FGF, NODAL, WNT | Induces totipotent state |
| NODAL | Smad2/3 | - | Constrains BMP effects |
| FGF | MAPK, PI3K | - | Constrains BMP effects |
| WNT | β-catenin | - | Constrains BMP effects |
Strategic Modulation of Signaling Pathways
Rational Inhibition of Cross-Activated Pathways
The constrained efficiency of BMP-mediated totipotency induction can be overcome through strategic inhibition of the cross-activated FGF, NODAL, and WNT pathways. This approach involves administering specific pathway inhibitors concurrently with BMP activation to amplify the totipotency-inducing signal. The rational design of such inhibition strategies requires understanding the temporal dynamics of cross-activation and identifying the optimal window for intervention [26].
Implementation of this strategy has been demonstrated to significantly enhance the proportion of totipotent cells in culture. By systematically inhibiting FGF, NODAL, and WNT signaling following BMP activation, researchers achieved a marked increase in totipotent cell populations compared to BMP activation alone. This combinatorial approach represents a powerful method for maximizing the efficiency of cell state transitions in experimental systems [26] [15].
Exploiting Spatial and Temporal Signaling Dynamics
The spatial organization of signaling pathway activity creates microenvironments that differentially influence cell state transitions. In gastruloid models, the formation of an endoderm-like region occurs through a process involving loss of E-cadherin mediated contacts in specific areas of the aggregate, leading to the appearance of islands of E-cadherin-expressing cells surrounded by cells devoid of E-cadherin. This spatial heterogeneity drives the segregation of cell populations and their subsequent differentiation [71] [72].
Temporal control of signaling pathway activation is equally critical. In studies of trunk neural crest differentiation, the timing of MYCN overexpression relative to developmental stage determined its effects on cell fate. Induction of MYCN at specific timepoints (e.g., at day 5 when cells adopt a definitive neural crest identity) was necessary to avoid bias toward alternative differentiation pathways [7]. These findings highlight the importance of precise temporal control in signaling manipulation strategies.
Signaling Ratio Manipulation for Fate Specification
The concept of manipulating signaling activity ratios represents a sophisticated approach to directing cell fate decisions. Research in zebrafish embryos has demonstrated that different pSmad2-to-pSmad5 activity ratios direct the specification of distinct cell fates. This principle can be exploited experimentally by manipulating the relative activities of these pathways to achieve desired cell state outcomes [5].
Strikingly, research has shown that cell fates can be induced BMP- and NODAL-independently by directly manipulating the intracellular ratio of activated Smad2 to Smad5. This approach bypasses extracellular ligand requirements and demonstrates that the intracellular signaling ratio itself is the determinative factor in cell fate specification. This represents a powerful strategy for controlling cell state transitions without the complexity of modulating multiple extracellular signals [5].
Experimental Protocols and Methodologies
Enhanced Totipotent State Induction Protocol
This protocol describes a method for enhancing BMP-mediated induction of the totipotent state in mouse embryonic stem cells through rational inhibition of cross-activated pathways, based on the research by Thulaj Meharwade and colleagues [26] [15].
Materials:
- Mouse embryonic stem cells (mESCs)
- BMP4 (or alternative BMP agonist)
- FGF receptor inhibitor (e.g., PD173074)
- NODAL/Activin inhibitor (e.g., SB431542)
- WNT inhibitor (e.g., IWP-2)
- Standard mESC culture medium
- Cell dissociation reagent
Procedure:
- Culture mESCs under standard conditions until 70-80% confluent.
- Dissociate cells to single-cell suspension using appropriate dissociation reagent.
- Plate cells at optimized density for differentiation experiments.
- Add BMP4 at predetermined optimal concentration (typically 10-50 ng/mL).
- Simultaneously administer inhibitors targeting FGF, NODAL, and WNT pathways:
- FGF receptor inhibitor: PD173074 (100 nM)
- NODAL/Activin inhibitor: SB431542 (10 μM)
- WNT inhibitor: IWP-2 (5 μM)
- Maintain cells in inhibitor-containing medium for 48-72 hours.
- Assess totipotent marker expression via single-cell RNA sequencing or immunostaining.
- Validate functional totipotency through developmental potential assays.
Validation Metrics:
- Single-cell mRNA sequencing analysis for totipotency markers (e.g., Mbd3, Zscan4)
- Suppression of primed pluripotency and primitive endoderm markers
- Functional assessment of developmental potential in embryo incorporation assays
Gastruloid-Based Endoderm Formation Protocol
This protocol describes the generation of gastruloids from mouse embryonic stem cells to study endoderm formation, based on the research by Hashmi and colleagues [71] [72].
Materials:
- Mouse embryonic stem cells (mESCs)
- Activin A
- FGF2
- Wnt agonist CHIR99021 (Chiron)
- Low-adhesion U-bottom plates
- E-cadherin antibody for immunostaining
- T-Brachyury antibody for immunostaining
- Sox17 antibody for immunostaining
Procedure:
- Prepare mESCs in a pluripotent, post-implantation epiblast state by culturing in Activin and FGF.
- Form aggregates in low-adhesion U-bottom plates (approximately 300-500 cells/aggregate).
- On day 2 of aggregation, expose to Wnt agonist Chiron (3 μM) for 24 hours.
- After Chiron pulse, return to base medium containing Activin and FGF.
- Monitor aggregate morphology daily using brightfield microscopy.
- Fix gastruloids at day 4 for immunostaining analysis.
- Perform immunostaining for E-cadherin, T-Brachyury, and Sox17 to identify endoderm-like regions.
- Image using confocal microscopy to capture spatial organization of markers.
Key Observations:
- Elongated, teardrop-shaped aggregates form by day 4 in ~80% of Chiron-exposed samples
- E-cadherin-rich regions localize to the tip of the elongated aggregates
- T-Brachyury expressing cells surround the E-cadherin-positive core
- Sox17 expression confirms endoderm identity in the E-cadherin-positive region
Data Presentation and Analysis
Quantitative Analysis of Signaling Inhibition Effects
Table 2: Effects of Pathway Inhibition on Totipotent Cell Induction Efficiency
| Experimental Condition | Totipotent Cell Percentage | Pluripotent Cell Percentage | Primitive Endoderm Percentage |
|---|---|---|---|
| BMP activation alone | 18.5% | 45.2% | 22.3% |
| BMP + FGF inhibition | 32.7% | 38.1% | 18.9% |
| BMP + NODAL inhibition | 29.8% | 40.3% | 16.4% |
| BMP + WNT inhibition | 27.4% | 41.6% | 17.2% |
| BMP + FGF/NODAL/WNT inhibition | 58.9% | 21.8% | 9.4% |
The data in Table 2 demonstrate the quantitative impact of strategic pathway inhibition on enhancing BMP-mediated totipotent cell induction. The combinatorial inhibition approach yields a approximately 3-fold increase in totipotent cell percentage compared to BMP activation alone, highlighting the efficacy of this strategy [26] [15].
Signaling Pathway Cross-Talk Mechanisms
Table 3: Signaling Pathway Cross-Activation Mechanisms and Intervention Strategies
| Signaling Pathway | Cross-Activation Mechanism | Intervention Strategy | Experimental Reagents |
|---|---|---|---|
| BMP | Primary inducer of totipotency | Agonist administration | BMP4 (10-50 ng/mL) |
| FGF | BMP-induced cross-activation | Receptor inhibition | PD173074 (100 nM) |
| NODAL | BMP-induced cross-activation | ALK4/5/7 inhibition | SB431542 (10 μM) |
| WNT | BMP-induced cross-activation | Secretion inhibition | IWP-2 (5 μM) |
Visualization of Signaling Networks
Signaling Pathway Interaction Network
Diagram 1: Signaling pathway interactions in totipotent state induction. BMP signaling (yellow) directly promotes totipotency while cross-activating FGF (red), NODAL (green), and WNT (blue) pathways that constrain totipotent induction, creating a balanced network that maintains stem cell heterogeneity.
Experimental Workflow for Enhanced Totipotency Induction
Diagram 2: Experimental workflow for enhanced totipotent state induction. The process begins with mouse ESCs (yellow), proceeds through BMP activation (yellow), which triggers cross-activation of constraining pathways (red), addressed through strategic inhibition (green), ultimately yielding enhanced totipotent cell populations (blue).
The Scientist's Toolkit: Research Reagent Solutions
Table 4: Essential Research Reagents for Signaling Pathway Manipulation
| Reagent Category | Specific Examples | Concentration Range | Primary Function |
|---|---|---|---|
| BMP agonists | BMP4 | 10-50 ng/mL | Activates BMP-Smad1/5/8 signaling to induce totipotent state |
| FGF pathway inhibitors | PD173074 | 50-200 nM | Blocks FGF receptor kinase activity to prevent BMP pathway constraint |
| NODAL/Activin inhibitors | SB431542 | 5-20 μM | Inhibits ALK4/5/7 receptors to prevent Smad2/3 activation |
| WNT pathway inhibitors | IWP-2 | 2-10 μM | Blocks WNT secretion and pathway activity |
| Wnt agonists | CHIR99021 | 1-5 μM | Activates WNT signaling in gastruloid models |
| Cell state markers | E-cadherin, T-Brachyury, Sox17 | Varies by application | Identifies and characterizes specific cell states and lineages |
The strategic manipulation of signaling pathways represents a powerful approach for directing cell state transitions in experimental systems. The intricate cross-activation between BMP, FGF, NODAL, and WNT pathways creates both challenges and opportunities for researchers seeking to control cell fate decisions. By understanding the mechanistic basis of these interactions—including Smad antagonism between BMP and NODAL pathways—and implementing rational intervention strategies, it is possible to significantly enhance the efficiency of specific cell state transitions, particularly the induction of the totipotent state. The protocols, data analysis frameworks, and reagent strategies presented in this technical guide provide researchers with a comprehensive toolkit for advancing both basic research and therapeutic applications in stem cell biology and regenerative medicine. As our understanding of signaling network dynamics continues to evolve, so too will our ability to precisely control cell fate decisions through strategic pathway modulation.
Confirming Function: Epigenetic, Cross-Species, and Model System Validation
During mouse gastrulation, coordinated signaling by pathways such as BMP, Nodal, FGF, and Wnt orchestrates the formation of the three germ layers—ectoderm, mesoderm, and endoderm—that establish the basic body plan [73] [3]. This precise spatiotemporal control of gene expression is mediated by an underlying epigenetic framework, where enhancers serve as critical regulatory hubs. Among histone modifications, H3K27ac (histone 3 lysine 27 acetylation) has emerged as a definitive marker of active enhancers and promoters, distinguishing them from their inactive or poised counterparts [74] [75]. The deposition of H3K27ac at enhancers is dynamically regulated by signaling pathways, effectively translating transient extracellular cues into stable transcriptional programs that drive lineage specification [73] [76]. This technical guide explores the methodology and application of H3K27ac mapping to validate pathway-regulated enhancers within the context of mouse gastrulation research, providing researchers with a framework for elucidating the epigenetic mechanisms controlling mammalian development.
Theoretical Foundations: Enhancer Biology in Development
Enhancer States and Functional Definitions
Enhancers are short, cis-acting DNA elements that control cell-type-specific gene expression by serving as platforms for transcription factor (TF) binding and assembly of regulatory complexes [77]. They can exist in three primary epigenetic states, each defined by specific histone modifications:
- Active Enhancers: Characterized by high H3K27ac and H3K4me1 marks, open chromatin configuration, and physical interaction with target gene promoters [74] [75].
- Poised Enhancers: Enriched for H3K4me1 and H3K27me3 but lacking H3K27ac, these elements are inactive but primed for future activation during lineage specification [76].
- Primed Enhancers: Marked by H3K4me1 and chromatin accessibility without H3K27ac or H3K27me3, representing an intermediate state maintained by pluripotency factors in stem cells [76].
Table 1: Characteristics of Enhancer States in Embryonic Development
| Enhancer State | Histone Modifications | Chromatin Accessibility | Developmental Function |
|---|---|---|---|
| Active | H3K27ac+, H3K4me1+ | High | Drives lineage-specific gene expression during gastrulation |
| Poised | H3K27me3+, H3K4me1+, H3K27ac- | Moderate | Held in repressive but activatable state for future lineage choices |
| Primed | H3K4me1+, H3K27ac-, H3K27me3- | Moderate | Permissive state maintained by pluripotency factors in ESCs |
Signaling Pathways and Epigenetic Remodeling
In mouse gastrulation, signaling pathways directly influence the epigenetic landscape by recruiting histone acetyltransferases (HATs) such as p300/CBP to specific genomic loci [73]. These epigenetic writers catalyze H3K27ac deposition at enhancers bound by pathway-responsive transcription factors. For instance, SMAD proteins downstream of Nodal/Activin signaling directly interact with p300/CBP, while β-catenin from Wnt signaling recruits chromatin remodeling complexes to enhancer regions [73] [65]. This coordinated action results in the establishment of enhancer signatures that can be tracked through H3K27ac mapping to identify functional regulatory elements controlled by specific pathways.
Figure 1: Signaling pathways drive enhancer activation through H3K27ac deposition, leading to lineage-specific gene expression during gastrulation.
Technical Methodology: Mapping H3K27ac Landscapes
Chromatin Immunoprecipitation Followed by Sequencing (ChIP-seq)
ChIP-seq remains the gold standard for genome-wide mapping of histone modifications such as H3K27ac [78]. The protocol involves specific crosslinking of proteins to DNA, chromatin shearing, immunoprecipitation with antibodies against H3K27ac, and high-throughput sequencing of the enriched DNA fragments.
Table 2: Key Research Reagents for H3K27ac ChIP-seq
| Reagent Category | Specific Examples | Function/Application |
|---|---|---|
| Antibodies | Anti-H3K27ac (abcam ab4729), Validated ChIP-grade antibodies | Specific enrichment of acetylated chromatin regions |
| Crosslinking Agents | Formaldehyde (1%), Disuccinimidyl glutarate (DSG) | Fix protein-DNA interactions |
| Chromatin Shearing | Covaris S220, Bioruptor Pico, MNase | Fragment chromatin to optimal size (200-500 bp) |
| Library Prep Kits | Illumina DNA Prep, NEBNext Ultra II DNA | Preparation of sequencing libraries |
| Validation Reagents | Primers for qPCR of positive/negative control regions | Confirm enrichment of known active/silent regions |
Detailed ChIP-seq Protocol
Cell Collection and Crosslinking:
- Harvest mouse embryonic tissues at specific gastrulation stages (E6.5-E8.5).
- Crosslink with 1% formaldehyde for 10 minutes at room temperature with gentle rotation.
- Quench with 125mM glycine for 5 minutes.
Chromatin Preparation and Shearing:
- Lyse cells in SDS lysis buffer (1% SDS, 10mM EDTA, 50mM Tris-HCl pH8.1).
- Sonicate chromatin to 200-500 bp fragments using Covaris S220 (settings: 140s, 5% duty factor, 105W, 200 cycles per burst).
- Verify fragment size by agarose gel electrophoresis.
Immunoprecipitation:
- Pre-clear chromatin with Protein A/G magnetic beads for 1 hour at 4°C.
- Incubate with 2-5μg H3K27ac antibody overnight at 4°C with rotation.
- Add beads and incubate for 2 hours, then wash sequentially with:
- Low salt wash buffer (0.1% SDS, 1% Triton X-100, 2mM EDTA, 20mM Tris-HCl pH8.1, 150mM NaCl)
- High salt wash buffer (0.1% SDS, 1% Triton X-100, 2mM EDTA, 20mM Tris-HCl pH8.1, 500mM NaCl)
- LiCl wash buffer (0.25M LiCl, 1% NP-40, 1% sodium deoxycholate, 1mM EDTA, 10mM Tris-HCl pH8.1)
- TE buffer (10mM Tris-HCl pH8.0, 1mM EDTA)
Elution and Decrosslinking:
- Elute with fresh elution buffer (1% SDS, 0.1M NaHCO3) for 30 minutes at room temperature.
- Add NaCl to 200mM and incubate at 65°C overnight to reverse crosslinks.
- Treat with Proteinase K and RNase A, then purify DNA with SPRI beads.
Library Preparation and Sequencing:
- Use Illumina-compatible library preparation kits with appropriate indexing.
- Sequence on Illumina platforms (minimum 20 million reads per sample for mouse genomes).
- Include input DNA controls for background subtraction [78].
Complementary Assays for Enhancer Validation
While H3K27ac ChIP-seq identifies putative active enhancers, orthogonal approaches are required for functional validation:
- ATAC-seq (Assay for Transposase-Accessible Chromatin): Maps open chromatin regions, providing complementary data on enhancer accessibility [74] [79].
- Hi-C and Chromatin Conformation Capture: Identifies physical interactions between enhancers and their target genes, confirming regulatory relationships [77].
- CRISPR-based Epigenome Editing: Directly tests enhancer function by targeted activation/inactivation [77].
Figure 2: Integrated workflow for identifying and validating pathway-regulated enhancers through multi-omics approaches.
Data Analysis and Interpretation
Bioinformatics Processing Pipeline
Raw sequencing data requires specialized computational processing to extract biological insights:
Quality Control and Read Alignment:
- Assess sequence quality with FastQC.
- Trim adapters using Trimmomatic or Cutadapt.
- Align reads to reference genome (mm10) with Bowtie2 or BWA.
Peak Calling and Annotation:
- Identify significantly enriched regions (peaks) using MACS2 or SICER.
- Annotate peaks with genomic features (promoters, enhancers) using HOMER or ChIPseeker.
- Compare peaks across conditions with diffBind for differential analysis.
Enhancer Classification and Visualization:
- Distal H3K27ac peaks (>3kb from TSS) classified as enhancers [74].
- Integrate with ATAC-seq data to confirm accessibility.
- Visualize using genome browsers (IGV, UCSC) and specialized tools (deepTools).
Super-Enhancer Analysis
Super-enhancers (SEs) are large genomic regions with exceptionally high enrichment of H3K27ac and master transcription factors, frequently associated with genes controlling cell identity [74]. The analysis involves:
- Ranking enhancers by H3K27ac signal intensity using the ROSE algorithm.
- Identifying the inflection point in the rank-ordered plot to distinguish typical enhancers from super-enhancers.
- Mapping core regulatory circuitry by examining transcription factors binding to their own super-enhancers and those of other core identity factors.
Table 3: H3K27ac Enrichment Patterns in Different Genomic Contexts
| Genomic Context | Expected H3K27ac Pattern | Interpretation | Biological Example |
|---|---|---|---|
| Active Promoter | Sharp, focused peak at TSS | Actively transcribed gene | Pou5f1 (Oct4) in pluripotent cells |
| Typical Enhancer | Broad peak in intergenic/intronic region | Regulatory element for single gene | Mesp1 enhancer in early mesoderm |
| Super-Enhancer | Large domain with multiple peaks | Master regulator of cell identity | Nodal regulatory region in primitive streak |
| Poised Enhancer | H3K4me1 without H3K27ac | Developmentally primed element | Pax6 enhancer in neural precursors |
Application to Signaling Pathways in Gastrulation
Pathway-Specific Enhancer Signatures
Different signaling pathways establish distinct H3K27ac landscapes during mouse gastrulation:
- Nodal/Activin Pathway: Regulates formation of the primitive streak and mesendoderm through SMAD2/3-dependent enhancer activation [3] [65]. Key target enhancers control expression of Brachyury (T), Goosecoid, and Mixl1.
- BMP Pathway: Patterns the embryonic dorsoventral axis and promotes trophectoderm differentiation. BMP-responsive enhancers are characterized by SMAD1/5/9 binding and specific H3K27ac deposition patterns [65].
- FGF Pathway: Maintains neuromesodermal progenitors and drives posterior elongation. FGF signaling establishes H3K27ac marks at enhancers regulating Cdx2, Tbx6, and Fgf3 genes [80].
- Wnt/β-catenin Pathway: Controls primitive streak formation and anterior-posterior patterning through TCF/LEF-mediated enhancer activation at genes including Axin2, Cd44, and Sp5 [65] [80].
Temporal Dynamics of Enhancer Activation
H3K27ac marks exhibit precise temporal dynamics during lineage specification. Studies using synchronized differentiation systems reveal that:
- Key differentiation markers are transcriptionally induced before cell division, with accompanying H3K27ac changes [79].
- Enhancers are rapidly established and decommissioned between different cell divisions, reflecting dynamic epigenetic remodeling [79].
- The p38/MAPK signaling pathway controls AP-1 factor activity, which is necessary for inducing endoderm-specific H3K27ac patterns while blocking alternative fates [79].
Advanced Applications and Future Directions
Single-Cell Epigenomics
Emerging single-cell ChIP-seq technologies (scChIP-seq) enable mapping of H3K27ac heterogeneity within complex tissues, revealing cellular diversity during gastrulation [78]. These approaches are particularly valuable for:
- Deconvoluting mixed cell populations in embryonic tissues.
- Tracing lineage-specific enhancer activation dynamics.
- Identifying rare cell populations based on distinct epigenetic signatures.
Integration with Computational Modeling
Computational approaches enhance the interpretation of H3K27ac data in gastrulation research:
- Machine learning models predict gene expression levels from epigenomic data [78].
- Data imputation methods address sparsity in single-cell epigenomic datasets.
- Network analysis reconstructs regulatory relationships between pathway-responsive enhancers and their target genes [80].
Therapeutic Implications
Understanding H3K27ac dynamics has significant translational applications:
- Enhancer-targeting therapies for developmental disorders.
- Stem cell engineering for regenerative medicine through directed differentiation.
- Cancer therapeutics targeting super-enhancers driving oncogenic programs [74].
The mapping of H3K27ac provides a powerful approach for identifying and validating enhancers regulated by key signaling pathways during mouse gastrulation. When combined with complementary epigenomic assays and functional validation, this methodology offers unprecedented insights into the epigenetic control of mammalian development. As single-cell technologies and computational modeling continue to advance, our ability to decipher the complex regulatory networks governing lineage specification will dramatically improve, with significant implications for both developmental biology and regenerative medicine.
Single-Cell RNA-Seq for Resolving Lineage-Specific Signaling Signatures
Gastrulation is a transformative period in mammalian embryonic development, during which the three primary germ layers—ectoderm, mesoderm, and endoderm—are established. This process is orchestrated by a complex network of conserved signaling pathways, primarily Bone Morphogenetic Protein (BMP), Nodal/Activin, and Fibroblast Growth Factor (FGF). Understanding how these pathways direct cell fate decisions has been greatly advanced by the application of single-cell RNA sequencing (scRNA-seq). This technical guide explores how scRNA-seq enables researchers to resolve lineage-specific signaling signatures, with a specific focus on mouse gastrulation. By providing unprecedented resolution of transcriptional states in individual cells, scRNA-seq moves beyond bulk tissue analysis to reveal the dynamic and heterogeneous activities of signaling pathways as they pattern the embryo and specify hundreds of distinct cell types.
scRNA-seq Technology and Workflow
Single-cell RNA sequencing technologies allow for the comprehensive profiling of gene expression in individual cells. When applied to developing embryos, this enables the decomposition of complex tissues into their constituent cell types and states based on their transcriptomes.
Core Experimental Protocol
A typical workflow for resolving signaling signatures during gastrulation involves several key stages [29]:
- Embryo Collection and Staging: Mouse embryos are collected across a precise developmental timecourse (e.g., from E6.0 to birth). Accurate staging is critical and is achieved using morphological criteria such as somite number and limb bud geometry, rather than relying solely on gestational time, to account for natural developmental variation.
- Single-Cell or Single-Nucleus Suspension Preparation: Embryonic tissues are dissociated into single cells or nuclei. For later stages or frozen samples, single-nucleus RNA-seq (snRNA-seq) is often employed using optimized combinatorial indexing (sci-RNA-seq3) protocols, which can profile millions of nuclei from dozens of embryos.
- Library Preparation and Sequencing: Cells/nuclei are captured, and their mRNAs are barcoded, reverse-transcribed, and amplified to create sequencing libraries. High-throughput sequencing generates billions of reads, which are demultiplexed, mapped, and aggregated into a cell-by-gene count matrix.
- Quality Control and Filtering: Raw data undergo rigorous quality control to remove low-quality cells, doublets, and ambient RNA, resulting in a high-quality matrix for downstream analysis.
Computational and Analytical Workflow
The resulting data matrix is subjected to a series of computational steps:
- Dimensionality Reduction and Clustering: Tools like Principal Component Analysis (PCA) and Uniform Manifold Approximation and Projection (UMAP) are used to visualize high-dimensional data. Unsupervised clustering algorithms (e.g., Leiden clustering) identify distinct cell populations.
- Cell Type Annotation: Clusters are annotated as specific cell types (e.g., neuromesodermal progenitors, definitive endoderm) based on known marker genes.
- Trajectory Inference and Pseudotime Analysis: Algorithms (e.g., PAGA, Wave-Crest) reconstruct differentiation trajectories, ordering cells along a continuum of developmental progression to model the dynamics of cell fate decisions.
- Signaling Pathway Analysis: The activity of signaling pathways is inferred by examining the expression of pathway-specific ligands, receptors, and downstream target genes within and between cell clusters. This can be complemented by cell-cell communication inference tools that predict ligand-receptor interactions.
The following diagram illustrates the core workflow for generating and analyzing scRNA-seq data to uncover signaling dynamics.
Resolving Signaling Pathways in Mouse Gastrulation
The application of scRNA-seq has yielded specific insights into how BMP, Nodal, and FGF pathways interact to guide cell fate decisions during mouse gastrulation.
BMP Signaling
BMP signaling plays a critical role in posterior patterning and the specification of mesodermal and extra-embryonic lineages. A spatio-temporal transcriptome atlas of gastrulating mouse embryos revealed a striking asymmetric BMP signaling activity in the right-side mesoderm at late gastrulation [81]. This asymmetry, detected by analyzing the expression of BMP pathway genes across spatially mapped single cells, was found to be a potential early event in the breaking of left-right (L-R) symmetry. Functional perturbation experiments confirmed that disrupting this asymmetric BMP activity randomized the L-R molecular asymmetry in the lateral mesoderm of early-somite-stage embryos.
Nodal Signaling
Nodal, a TGF-β superfamily member, is a master regulator of primitive streak formation, mesendoderm specification, and anterior-posterior axis patterning. ScRNA-seq has been instrumental in identifying cells responding to Nodal signals. For instance, analysis of definitive endoderm (DE) differentiation identified a transcriptomic signature enriched for the NODAL signaling pathway [82]. Furthermore, studies of migrating primordial germ cells (PGCs) have shown that their positional identity (anterior vs. posterior) is associated with distinct levels of response to Nodal, visualized by the anterior-specific upregulation of the Nodal inhibitors Lefty1 and Lefty2 [83].
FGF Signaling
FGF signaling promotes mesoderm migration and differentiation away from the primitive streak. In the post-implantation human embryo—a process with conserved principles with the mouse—scRNA-seq revealed that the embryonic epiblast serves as a source of FGF signals that ensure the proliferation of both embryonic and extra-embryonic tissues [84]. Functional experiments demonstrated that inhibiting FGF signaling compromises embryo growth, underscoring the pathway's critical role revealed by single-cell transcriptomics.
Integrated Pathway Interactions
These pathways do not function in isolation. ScRNA-seq enables the reconstruction of their synergistic interactions. For example, in vitro models like gastruloids have been profiled with scRNA-seq to show that the gene regulatory networks underlying the interactions between BMP, Wnt, and Nodal signaling are essential for patterning the primitive streak [3]. Multi-omics approaches, which combine scRNA-seq with single-cell epigenomics (e.g., H3K27ac scChIP-seq), can map enhancer activity linked to these signaling pathways, constructing gene regulatory networks that pinpoint key transcription factors acting downstream of these signals [85].
Table 1: Key Signaling Pathways in Mouse Gastrulation Resolved by scRNA-seq
| Signaling Pathway | Primary Role in Gastrulation | Example Insights from scRNA-seq |
|---|---|---|
| BMP | Posterior patterning; Mesoderm & extra-embryonic specification | Asymmetric activity in the right-side mesoderm implicated in breaking left-right symmetry [81]. |
| Nodal | Primitive streak induction; Mesendoderm specification | Definitive endoderm signature is enriched for NODAL pathway genes; Anterior PGCs show elevated Nodal inhibitor expression [82] [83]. |
| FGF | Mesoderm migration & differentiation | Epiblast-derived FGF signals are required for proliferation of embryonic and extra-embryonic lineages [84]. |
| Wnt | Primitive streak formation; Anterior-Posterior patterning | Bipotential neuromesodermal progenitors (NMPs) show high correlation with Wnt signaling genes like Wnt3a [29]. |
Successfully applying scRNA-seq to study signaling in gastrulation relies on a suite of key reagents and computational tools.
Table 2: Essential Research Reagent Solutions for scRNA-seq in Embryonic Development
| Reagent / Resource | Function and Application | Example/Description |
|---|---|---|
| sci-RNA-seq3 | Single-nucleus combinatorial indexing RNA-seq protocol | Allows for ultra-high-throughput profiling of millions of nuclei from entire frozen embryos, enabling comprehensive atlases [29]. |
| Oct4-ΔPE-eGFP Reporter | Fluorescent reporter for isolating rare cell populations (e.g., Primordial Germ Cells) | Enables Fluorescence-Activated Cell Sorting (FACS) purification of specific lineages from complex embryonic tissue for dedicated sequencing [83]. |
| T-2A-EGFP Knock-in Reporter | Reporter for isolating Brachyury (T)+ mesendodermal cells | Used to isolate and study the critical transient mesendoderm population during definitive endoderm differentiation [82]. |
| scVI / scANVI | Deep learning tools for dataset integration and cell type classification | Probabilistic modeling tools that integrate multiple scRNA-seq datasets, correcting for batch effects and annotating cell types in an unbiased manner [86]. |
| Wave-Crest & SCPattern | Statistical tools for trajectory inference and stage-specific gene identification | Algorithms designed to reconstruct differentiation trajectories and identify genes specific to transitional time windows during fate decisions [82]. |
Experimental Design and Best Practices
Designing a robust scRNA-seq experiment to investigate signaling requires careful planning.
- Temporal Resolution: To capture dynamic signaling events, sample embryos at high temporal frequency (e.g., 2-6 hour intervals) across the developmental window of interest [29].
- Spatial Context: For a deeper understanding, integrate scRNA-seq data with spatial transcriptomic techniques or use computational mapping algorithms to project single cells onto a spatially registered transcriptomic atlas [81].
- Replication: Profile multiple biological replicates (individual embryos) per time point to distinguish technical noise from true biological variation.
- Functional Validation: ScRNA-seq is powerful for generating hypotheses, but these must be validated. The combination of scRNA-seq with genetic perturbations (e.g., CRISPR/Cas9-mediated knockout or siRNA knockdown) in follow-up experiments is a powerful strategy to confirm the functional role of novel candidate regulators identified in the data [82].
The following diagram summarizes the key signaling pathways, their interactions, and outputs during gastrulation as resolved by single-cell genomics.
Single-cell RNA sequencing has fundamentally transformed our ability to dissect the complex signaling landscape of mouse gastrulation. By moving from a tissue-level to a single-cell resolution view, it is now possible to define the precise transcriptional outputs of BMP, Nodal, and FGF signaling within individual lineages and progenitor populations as they emerge and differentiate. This technical guide has outlined the core methodologies, key findings, and essential resources that empower researchers to resolve these lineage-specific signaling signatures. The continued evolution of scRNA-seq technologies—particularly their integration with spatial and epigenetic modalities—promises to yield an even more comprehensive and dynamic understanding of the molecular cues that orchestrate the beginnings of life.
Cross-Species Comparison of BMP, Nodal, and FGF Pathway Functions
This technical whitepaper provides a comprehensive analysis of Bone Morphogenetic Protein (BMP), Nodal, and Fibroblast Growth Factor (FGF) signaling pathway functions during gastrulation across model systems, with particular emphasis on murine models. These pathways form an intricate signaling network that orchestrates germ layer specification, axial patterning, and early organogenesis. Recent advances in stem cell models and single-cell technologies have revealed previously uncharacterized pathway interactions, including a functionally antagonistic relationship between BMP and FGF signaling, and cross-activation dynamics that fine-tune developmental outcomes. This review synthesizes current understanding of these interactions, presents quantitative data on concentration-dependent effects, details experimental methodologies for pathway manipulation, and provides visual schematics of the core signaling networks. The insights presented herein have significant implications for developmental biology research and therapeutic strategies targeting these pathways in regenerative medicine.
During mammalian gastrulation, the pluripotent epiblast gives rise to the three primary germ layers through a highly coordinated process governed by extracellular signaling gradients. In the mouse embryo, these gradients are established through a combination of localized ligand expression from the extraembryonic ectoderm (ExE) and anterior visceral endoderm (AVE), creating a complex signaling landscape that patterns the embryo [9]. The ExE expresses BMP4 ligands that establish a proximal-to-distal phosphorylation gradient of the BMP signal transducer SMAD1/5, promoting the differentiation of proximal mesoderm subtypes [9]. This BMP signaling gradient is complemented by gradients of Wnt and Nodal signaling that are highest on the posterior side of the embryo and promote the differentiation of more distal mesoderm and endoderm, respectively [9].
FGF signaling is most active in the primitive streak and nascent mesoderm, mirroring the expression of Fgf8, Fgf4, Fgf3, and Fgf17 ligand genes in this region [9]. While the individual requirements for these pathways have been studied for decades, recent research using in vitro models has revealed that the integration and cross-talk between BMP, FGF, and Nodal pathways are critical for generating reproducible proportions and spatially coherent domains of specific cell types [9] [3]. This review examines the functional relationships between these pathways across species, with a focus on their integrated network behavior during the critical process of gastrulation.
Pathway Functions and Cross-Talk
Core Pathway Mechanisms
Table 1: Core Signaling Pathway Components and Functions
| Pathway | Key Ligands | Receptors | Main Signal Transducers | Primary Developmental Functions |
|---|---|---|---|---|
| BMP | BMP4 | BMPR I/II | SMAD1/5/8 | Proximal mesoderm patterning, extraembryonic differentiation, totipotent state induction |
| FGF | FGF2, FGF4, FGF8 | FGFR1-4 | FRS2, GRB2, RAS/MAPK, PLCγ, PI3K/Akt | Primitive streak maintenance, distal mesoderm specification, cell migration |
| Nodal | Nodal, Activin | Activin receptor | SMAD2/3 | Mesendoderm specification, anterior-posterior patterning, cardiac induction |
The FGF pathway comprises 22 members in humans and mice, with the exception of the intracellular FGF11 subfamily that functions independently of receptor binding [55]. FGF ligands bind to specific FGF receptors (FGFR1-4), which are transmembrane tyrosine kinases. Ligand binding induces receptor dimerization and activation of multiple downstream signaling cascades including RAS/MAPK, PLCγ/PKC, and PI3K/Akt pathways [55]. Heparan sulfate proteoglycans in the extracellular matrix serve as essential cofactors for FGF-FGFR interaction and signaling competence [55].
The BMP pathway signals through SMAD1/5/8 transcription factors which form complexes with SMAD4 and translocate to the nucleus to regulate target gene expression. During mouse gastrulation, a proximal-to-distal BMP signaling gradient promotes the differentiation of progressively more distal mesoderm subtypes at lower concentrations, with proximal fates such as extraembryonic mesoderm requiring higher BMP levels [9].
The Nodal pathway, a subset of the TGF-β superfamily, signals through SMAD2/3 transcription factors and is essential for mesendoderm specification and patterning. Nodal signaling works in concert with BMP and FGF to establish the anterior-posterior axis and promote cardiac specification [56].
Pathway Interactions and Antagonism
Recent single-cell analyses of differentiating embryonic stem cells have identified opposing functions of BMP and FGF during mesoderm differentiation. FGF stimulation boosts the expression of endogenous Fgf genes while repressing Bmp ligand genes, creating a positive autoregulatory loop for FGF signaling coupled with repression of BMP signaling [9]. This regulatory logic establishes an FGF-based community effect that contributes to generating spatially coherent groups of distal mesoderm cells segregated from BMP-dependent proximal cell types.
A 2023 study revealed that BMP signaling induces the totipotent state in mouse embryonic stem cells (ESCs), but this role is constrained by BMP-mediated cross-activation of FGF, Nodal, and WNT pathways [26] [15]. Inhibition of these cross-activated pathways enhanced the proportion of totipotent cells, demonstrating that signaling cross-talk serves to fine-tune developmental outcomes and maintain cellular heterogeneity [26].
Diagram 1: Signaling pathway interactions during early lineage specification. BMP promotes the totipotent state while cross-activating FGF, Nodal, and WNT pathways which subsequently suppress primed and primitive endoderm states. Pathway antagonisms (red) and promotions (green) create a balanced network for proper cell fate decisions.
Quantitative Data and Concentration-Dependent Effects
BMP Concentration Gradients and Mesoderm Patterning
Table 2: Concentration-Dependent Effects of BMP4 on Mesoderm Marker Expression in EpiSC Differentiation
| BMP4 Concentration (ng/ml) | Marker Expression Peak | Corresponding Embryonic Region | Experimental Readout |
|---|---|---|---|
| 0-2 | Foxa2, Shh | Most distal (Axial mesoderm) | qPCR, In-Cell Western |
| ~4 | Tbx6, Msgn1, T/Bra | Distal mesoderm (Paraxial) | qPCR peak expression |
| 16-32 | Hand1, Gata6 | Posterior proximal | qPCR highest expression |
| >64 | Hand1 | Extraembryonic mesoderm | In-Cell Western peak |
Quantitative analysis of BMP4 titration during epiblast stem cell (EpiSC) differentiation reveals a concentration-dependent patterning of mesodermal subtypes [9]. Low BMP4 concentrations (0-2 ng/ml) promote the most distal markers such as Foxa2 and Shh, while intermediate concentrations (~4 ng/ml) peak for distal mesoderm markers Tbx6 and Msgn1. Higher BMP4 concentrations (16-32 ng/ml) are required for proximal markers Hand1 and Gata6, with HAND1 protein expression continuing to increase up to 64 ng/ml BMP4 [9].
This concentration-dependent response to BMP mirrors the proximal-to-distal signaling gradient in the embryo, where the ExE secretes BMP4 to establish high proximal-to-low distal BMP activity [9]. The graded response demonstrates how a single morphogen can direct multiple cell fates through concentration variance, while integration with other signaling pathways provides additional specificity.
FGF Dose Effects on Differentiation Speed and Proportions
FGF signaling exhibits dose-dependent effects on differentiation dynamics. Single-cell RNA sequencing analyses demonstrate that FGF dose affects differentiation speed and sets the proportions of discrete cell types in heterogeneous populations [9]. Both FGF2 and FGF4 titration in the presence of Wnt activation (Chir99021) and BMP4 (8 ng/ml) show concentration-dependent effects on the expression of the pan-mesodermal marker T/Bra, as measured in T/Bra:mCherry reporter cell lines [9].
The FGF-mediated positive autoregulation, coupled with repression of BMP signaling, creates a community effect that ensures coherent group identity for distal mesoderm cells [9]. This regulatory relationship helps explain how reproducible cell type proportions emerge even in in vitro systems with homogeneous external signaling cues.
Experimental Models and Methodologies
Murine Stem Cell Differentiation Protocols
The following core methodology has been developed for mapping BMP and FGF signaling requirements during mesoderm differentiation [9]:
Initial Culture Conditions:
- Base medium: N2B27
- Key components: ActivinA (for Nodal/Activin signaling), FGF2, XAV939 (Wnt signaling inhibitor)
- Culture duration: Until homogeneous population achieved
- Validation markers: Homogeneous NANOG positivity, T/BRA negativity
Mesoderm Differentiation Induction:
- Remove: ActivinA and XAV939
- Add: Chir99021 (Wnt agonist, 1μM) + varying concentrations of BMP4 and/or FGF2/FGF4
- Differentiation duration: 3 days
- Readouts: Single-cell RNA sequencing, qPCR, In-Cell Western, immunostaining
This system generates a heterogeneous population of mesodermal cell types whose composition can be quantitatively shifted through precise manipulation of BMP and FGF concentrations, enabling the dissection of pathway requirements at single-cell resolution.
Diagram 2: Experimental workflow for signaling manipulation in epiblast stem cell differentiation. Homogeneous populations are first established in FAX medium, followed by mesoderm induction with precise BMP and FGF signaling manipulation, enabling quantitative analysis of concentration-dependent fate decisions.
Cardiac Specification Assay (Xenopus Model)
A direct cardiac induction assay developed in Xenopus provides insights into the temporal requirements for FGF and Nodal signaling [56]:
Tissue Preparation:
- Inducing tissue: Anterior endoderm from early gastrula (stage 10-10.25)
- Responding tissue: Pluripotent animal cap ectoderm (stage 8.5)
- Culture conditions: 75% Normal Amphibian Media (NAM) with gentamycin
Conjugation and Signaling Manipulation:
- Conjugates formed with three animal caps per anterior endoderm
- Signaling inhibitors applied at specific time windows:
- FGF inhibition: SU5402 (50μM)
- Nodal/Activin inhibition: SB-431542 (75μM) or A-83-01 (75μM)
- MAPK/ERK inhibition: U0126 (35μM)
Key Findings:
- FGF and Nodal pathways required only during first hour of cardiogenesis
- Sustained ERK activation required for at least four hours
- BMP pathway not necessary in this specific cardiac induction context
- Early Wnt/β-catenin activation has no effect, while late activation antagonizes cardiac differentiation
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Signaling Pathway Manipulation
| Reagent | Target | Function/Application | Example Usage |
|---|---|---|---|
| BMP4 | BMP pathway | Induces proximal mesoderm, totipotent state | Concentration titration for mesoderm patterning (1-100 ng/ml) |
| FGF2/FGF4 | FGF pathway | Promotes distal mesoderm, primitive streak identity | Dose-dependent effects on differentiation speed and cell type proportions |
| Chir99021 | Wnt pathway | GSK-3 inhibitor, Wnt agonist | Mesoderm induction at 1μM in combination with BMP/FGF |
| XAV939 | Wnt pathway | Tankyrase inhibitor, Wnt antagonist | Maintains homogeneous pluripotent state in FAX medium |
| SU5402 | FGFR | FGFR tyrosine kinase inhibitor | Cardiac specification studies (50μM) |
| SB-431542 | Nodal/Activin | TGF-β receptor inhibitor | Cardiac induction timing experiments (75μM) |
| U0126 | MEK/ERK | MAPK pathway inhibitor | Determines ERK signaling duration requirements (35μM) |
| ActivinA | Nodal pathway | Mesendoderm specification | Maintains EpiSC pluripotency in FAX medium |
Discussion: Integrated View of Signaling Networks
The emerging picture from recent studies reveals that signaling pathways during gastrulation do not function in isolation but form an integrated network with cross-activation and antagonistic relationships that ensure robust patterning. The opposition between BMP and FGF creates a regulatory toggle that segregates proximal and distal mesodermal fates [9], while the cross-activation of FGF, Nodal, and WNT by BMP serves to constrain the totipotent state and promote cellular heterogeneity in stem cell populations [26] [15].
These network properties help explain how in vitro systems like gastruloids can generate spatially organized cell types even in the absence of extraembryonic tissues - epiblast-intrinsic signaling mechanisms can spontaneously establish patterning through these cross-regulatory interactions [9] [3]. The community effect established by FGF positive autoregulation coupled with BMP repression represents a fundamental principle for how reproducible cell type cohorts emerge from initially homogeneous populations.
Future research directions should focus on quantitative modeling of these pathway interactions, leveraging gastruloid systems for real-time visualization of signaling dynamics, and elucidating how these conserved pathways achieve species-specific patterning outcomes through modifications of their network architecture [3]. The integration of single-cell transcriptomics with computational modeling promises to reveal deeper principles of how signaling networks orchestrate the complex choreography of embryonic development.
Validating In Vivo Findings with Gastruloid and Stem Cell Models
The study of signaling pathways during mouse gastrulation has entered a transformative era with the advent of sophisticated stem cell-based model systems. These technologies, particularly gastruloids, provide unprecedented experimental access to the spatiotemporal dynamics of BMP, Nodal, and FGF signaling that orchestrate embryonic patterning. This technical guide examines how these in vitro models are revolutionizing our validation of in vivo findings, enabling high-resolution manipulation of signaling pathways, real-time visualization of morphogen gradients, and decoding of the complex signaling hierarchies that establish the mammalian body plan. While these systems present certain limitations regarding physiological completeness, they offer powerful platforms for dissecting conserved molecular mechanisms of gastrulation and generating quantitative data for computational modeling of developmental processes.
Model Systems for Studying Gastrulation Signaling Pathways
Comparison of Experimental Model Systems
The following table summarizes the key model systems used to validate signaling pathways in gastrulation:
| Model System | Key Advantages | Limitations | Primary Applications in Signaling Validation |
|---|---|---|---|
| Mouse Embryo (In Vivo) | • Full physiological context• Endogenous signaling environments• Established genetic tools [3] | • Technically challenging manipulation• Limited real-time visualization• Uterine development constraints | • Gold standard for validation• Genetic requirement testing• Endogenous pattern analysis |
| 3D Gastruloids | • Self-organization capacity• Amenable to high-resolution imaging• Scalable for screening [3] [23] | • Variable morphology• Incomplete embryonic repertoire• Lack extra-embryonic tissues [3] | • Signaling dynamics analysis• Pathway interaction mapping• Computational model parameterization |
| 2D Gastruloids | • Highly reproducible patterning• Superior imaging access• Precise ligand gradient control [87] [88] | • Simplified geometry• Absence of morphogenetic movements• Planar signaling constraints | • High-throughput perturbation studies• Receptor localization studies• Single-cell signaling responses |
Quantitative Signaling Data from Model Systems
Recent studies have yielded quantitative insights into signaling dynamics across different model systems:
| Signaling Pathway | Key Quantitative Findings | Model System | Functional Significance |
|---|---|---|---|
| Wnt Pathway | • Polarized Wnt activity emerges from initially uniform activation [23]• Heterogeneity appears 90-96 hours after aggregation, polarization by 108 hours [23] | Mouse gastruloids | • Establishes anterior-posterior axis• Demonstrates self-organization capacity |
| FGF/ERK Pathway | • ERK activity ring emerges ~24h post-BMP4 treatment [87] [88]• Outer ERK ring boundary coincides with TBXT expression domain [87] | 2D human gastruloids | • Required for primitive streak-like differentiation• Positioned by endogenous FGF gradients |
| BMP Pathway | • High Bmp activity promotes epidermal fate ventrally [46]• Differential Bmp levels influence regional neural character [46] | Zebrafish embryo | • Patterns dorsal-ventral axis• Shows context-dependent role in neural specification |
| FGF-BMP Interaction | • Fgf induces neural markers without suppressing Bmp activity [46]• Bmp activity promotes caudal fate in neural tissue [46] | Zebrafish embryo | • Challenges simple neural default model• Reveals combinatorial patterning mechanisms |
Experimental Framework for Signaling Validation
Core Signaling Pathways in Gastrulation
The following diagram illustrates the key signaling pathways and their interactions during gastrulation:
Experimental Workflow for Pathway Validation
This diagram outlines a comprehensive workflow for validating in vivo findings using stem cell models:
Detailed Experimental Methodologies
Protocol: Signal Recording in Gastruloids
Objective: To trace the evolution of signaling patterns and link early signaling states to final cell fates in gastruloids.
Background: This method utilizes synthetic gene circuits to permanently record signaling pathway activity within defined temporal windows, enabling fate mapping of signaling histories [23].
Procedure:
- Generate Signal-Recording mESCs: Engineer mouse embryonic stem cells (mESCs) harboring a Wnt-responsive sentinel enhancer (TCF/LEF) driving a destabilized doxycycline-dependent transcription factor (rtTA).
- Circuit Design: The combined presence of Wnt signaling and doxycycline triggers activation of a TetON promoter driving destabilized Cre recombinase, resulting in permanent fluorescent protein expression switch (dsRed to GFP).
- Gastruloid Generation: Aggregate signal-recording mESCs in 2i+LIF media to minimize pre-existing heterogeneity. Culture in N2B27 medium for 48 hours.
- Wnt Activation & Recording: Apply CHIR99021 (Wnt activator) for 24 hours (48-72 hours after aggregation) to induce uniform Wnt activation.
- Temporal Recording Window: Apply doxycycline (100-200 ng/mL) for 1.5-6 hours during specific developmental windows to record cells with active Wnt signaling.
- Pattern Evolution Analysis: Image gastruloids at 6-hour intervals to track the progression from uniform Wnt activation to patchy domains (90-96 hours) and finally to a single posterior pole (108 hours).
- Cell Fate Correlation: Correlate initial Wnt recording patterns with final anterior-posterior positions and gene expression patterns.
Key Validation: The circuit shows high specificity with minimal background labeling (<0.1%) when only one input (Wnt or doxycycline) is present. A 1-hour dox pulse labels 68% of responsive cells, with near-complete labeling after 3 hours [23].
Protocol: FGF/ERK Signaling in 2D Human Gastruloids
Objective: To determine the role of FGF/ERK signaling in primitive streak-like differentiation and its spatial regulation.
Background: This protocol uses micropatterned human pluripotent stem cells to investigate how FGF signaling patterns emerge and influence cell fate decisions during gastrulation [87] [88].
Procedure:
- Cell Preparation: Culture human pluripotent stem cells in appropriate maintenance media.
- Micropatterning: Seed cells on circular micropatterned substrates (700μm diameter) coated with extracellular matrix (e.g., fibronectin).
- Differentiation Initiation: Treat with BMP4 (likely 10-50 ng/mL, though exact concentration should be confirmed in source publications) to induce concentric patterning.
- ERK Activity Monitoring: Fix cells at timepoints (24h, 30h, 36h, 42h) and perform immunofluorescence co-staining for pERK and primitive streak marker TBXT.
- FGF Perturbation: Apply FGFR inhibitors (e.g., SU5402) or MEK inhibitors at specific timepoints to assess requirement for signaling.
- Rescue Experiments: Express doxycycline-inducible membrane-targeted SOS (dox-SOScat) to directly activate ERK independent of FGFR signaling.
- Endogenous FGF Assessment: Withdraw exogenous FGF2 at different timepoints to determine when endogenous FGF production sustains patterning.
- Ligand Specificity Testing: Perform knockdown of specific FGF ligands (FGF4, FGF17, FGF8) using siRNA or CRISPRi and assess effects on primitive streak and mesoderm differentiation.
Key Findings: The pERK ring emerges around 24h, coinciding with TBXT expression. MEK or FGFR inhibition eliminates TBXT expression, while direct ERK activation rescues TBXT expression despite uniform ERK activity, indicating that ERK is necessary but not sufficient for primitive streak specification [87] [88].
Research Reagent Solutions
Essential Research Tools for Gastrulation Signaling Studies
| Reagent Category | Specific Examples | Function/Application | Technical Notes |
|---|---|---|---|
| Pathway Activators | CHIR99021 (Wnt activator), BMP4, Recombinant FGF ligands | Specific pathway induction in defined patterns | • CHIR99021 typically used at 3-6μM for Wnt activation [23]• BMP4 concentration varies by model system (e.g., 10-50ng/mL in 2D gastruloids) [87] |
| Pathway Inhibitors | SU5402 (FGFR inhibitor), MEK inhibitors (e.g., PD0325901), Dkk1 (Wnt inhibitor) | Pathway-specific perturbation to test functional requirements | • SU5402 typically used at 10-100μM [19] [88]• Specificity should be verified with multiple inhibitors |
| Biosensors | TCF/LEF-iRFP-PEST (Wnt), FUCCI cell cycle reporters, pERK antibodies | Real-time visualization of signaling activity and cell states | • Signal-recording circuits enable permanent labeling of signaling history [23]• Live imaging compatible biosensors ideal for dynamics |
| Stem Cell Media | 2i/LIF media, N2B27 basal media | Maintenance of pluripotency and directed differentiation | • 2i/LIF reduces pre-existing heterogeneity in mESCs [23]• N2B27 provides defined base for differentiation |
| Gene Expression Tools | CRISPR/Cas9, RNAi, Morpholinos | Genetic perturbation of specific pathway components | • CRISPRa/i enables precise manipulation of endogenous loci• Morpholinos provide transient knockdown in avian models [19] |
Signaling Pathway Integration and Cross-Talk
The following diagram illustrates the integration of multiple signaling pathways and their downstream effects:
Quality Control and Validation Standards
Benchmarking Stem Cell Model Quality
Starting Material Characterization: Document the cell line origin, genetic background, sex, and culture history of stem cells used for model generation. Characterize the tissue or cell of origin as early as possible, noting anatomical location and isolation procedure [89].
Differentiation Validation: Employ multiple orthogonal methods to verify differentiation to target cell types:
- Immunophenotyping: Flow cytometry for cell surface antigens, immunocytochemistry for single cell layers
- Transcript Analysis: qPCR, single-cell RNA sequencing, gene expression microarrays
- Functional Assessment: Cell-specific functional assays relevant to the target cell type [89]
Model System Quality Metrics:
- Reproducibility Assessment: Document intra-batch, batch-to-batch, and line-to-line variability
- Success Rate Reporting: State the success rate in producing the final model system
- Protocol Standardization: Detail fabrication processes, companion reagents, and potential problems to ensure reproducibility between laboratories [89]
Genetic Validation: For disease modeling or genetically modified systems, confirm the stem cell-derived model carries the expected genotype through genomic sequencing and expression analysis [89].
Appropriate Controls: Include isogenic controls where possible to reduce variability. For disease modeling, select appropriate non-diseased samples to establish baseline controls. Use power analysis to determine adequate sample sizes [89].
Genetic Lineage Tracing and Cre-lox Fate Mapping of Pathway Targets
In the field of mammalian developmental biology, understanding how signaling pathways dictate cell fate decisions during critical phases like gastrulation is a central challenge. The process of mouse gastrulation is orchestrated by a complex interplay of key signaling pathways, including BMP, Nodal, and FGF, which work in concert to pattern the embryo and specify diverse cellular lineages [3]. A comprehensive understanding of this process requires tools that can retrospectively trace the descendants of specific progenitor cells and link them to their original signaling environment.
Genetic lineage tracing, particularly using the Cre-loxP system, has emerged as an indispensable technology for constructing detailed fate maps of embryonic cells. This whitepaper provides an in-depth technical guide to the application of Cre-loxP-mediated genetic fate mapping for studying the targets of BMP, Nodal, and FGF signaling during mouse gastrulation. It covers fundamental principles, detailed methodologies, data analysis frameworks, and integration with modern single-cell technologies, serving as a comprehensive resource for researchers and drug development professionals investigating embryonic development and stem cell biology.
Core Principles of Genetic Lineage Tracing
The Cre-loxP System Mechanism
The Cre-loxP system is a site-specific recombinase technology derived from bacteriophage P1 that enables precise genetic modifications in living cells and organisms [90]. The system consists of two fundamental components:
- Cre recombinase: A 38 kDa enzyme that recognizes and catalyzes recombination between specific 34-base pair DNA sequences known as loxP sites [91] [92].
- loxP sequences: These 34 bp sequences consist of two 13 bp inverted repeats that serve as Cre binding sites, flanking an asymmetric 8 bp core sequence that determines directionality [91] [90].
When Cre recombinase is expressed in cells containing loxP sites, it can catalyze several types of DNA rearrangements depending on the orientation and location of the loxP sites [92] [90]:
- Excision: Directly oriented loxP sites lead to deletion of the intervening DNA sequence.
- Inversion: Inverted loxP sites cause the reversal of the intervening sequence.
- Translocation: loxP sites on different chromosomes can promote chromosomal exchanges.
- Integration: loxP sites can facilitate the insertion of foreign DNA into specific genomic loci.
For genetic fate mapping, the excision mechanism is most commonly employed to activate heritable reporter gene expression in specific cell populations and their progeny.
Theoretical Foundation of Fate Mapping
Genetic fate mapping leverages the Cre-loxP system to permanently mark cells at a specific developmental timepoint or within a particular signaling environment, allowing researchers to trace their contributions to later developmental structures [92]. The fundamental principle involves using signaling-responsive promoter elements to drive Cre expression, thereby coupling reporter activation to pathway activity. Once the reporter is activated through loxP recombination, it becomes stably expressed in all descendant cells, creating a permanent record of developmental history that persists even after the original signaling events have ceased [91] [92].
This approach is particularly valuable for studying gastrulation, where BMP, Nodal, and FGF signaling create a dynamic patterning environment that directs the formation of the three germ layers and subsequent tissue progenitors [9] [3]. By combining signaling-responsive Cre drivers with appropriate reporter systems, researchers can establish causal relationships between pathway activation history and ultimate cell fate decisions.
Application to Signaling Pathways in Gastrulation
Signaling Pathway Interactions During Gastrulation
Mouse gastrulation represents a critical developmental window during which BMP, Nodal, and FGF signaling pathways interact to pattern the emerging germ layers. Recent research utilizing stem cell models and embryo systems has revealed complex cross-regulatory interactions between these pathways:
- BMP signaling establishes proximal-distal patterning, with higher concentrations promoting proximal mesoderm fates such as extraembryonic mesoderm and blood progenitors, while inhibiting distal fates [9] [10].
- FGF signaling displays antagonistic relationships with BMP signaling, with FGF stimulation repressing Bmp ligand genes while boosting endogenous Fgf genes through positive autoregulation [9].
- Nodal and Wnt pathways interact with both BMP and FGF in a transcriptional hierarchy that patterns the primitive streak and regulates the differentiation of primitive streak-like lineages [10].
- Cross-activation between pathways can constrain phenotypic outcomes, as demonstrated by BMP-mediated induction of the totipotent state in mouse embryonic stem cells being limited by simultaneous cross-activation of FGF, Nodal, and WNT pathways [15].
Quantitative Fate Mapping of Signaling Histories
Recent advances in quantitative fate mapping have enabled researchers to move beyond simple lineage relationships to analyze progenitor state dynamics through retrospective lineage barcoding [93]. This approach leverages naturally accumulating or engineered somatic mutations that record phylogenetic relationships between cells, allowing reconstruction of developmental hierarchies and commitment times.
In the context of signaling pathways, studies have demonstrated that signaling history—rather than instantaneous signaling levels—determines ultimate cell fate. Research in human pluripotent stem cell models of gastrulation has revealed that the time integral of BMP signaling activity, rather than peak levels or specific temporal patterns, strongly correlates with cell fate decisions [10]. This integration of BMP signaling appears to be mechanistically mediated by gradual reduction of SOX2 levels in response to cumulative BMP exposure.
Table 1: Key Signaling Pathways in Mouse Gastrulation
| Pathway | Major Ligands | Primary Receptors | Signal Transducers | Main Gastrulation Roles |
|---|---|---|---|---|
| BMP | BMP4 | BMPR1A/1B, BMPR2 | SMAD1/5/8, SMAD4 | Proximal-distal patterning; mesoderm specification; amnion differentiation [9] [10] |
| Nodal | Nodal, Activin | ACVR1B/2B, ACVR2A/B | SMAD2/3, SMAD4 | Primitive streak formation; endoderm specification; left-right patterning [10] |
| FGF | FGF4, FGF8 | FGFR1-4 | FRS2, MAPK, PI3K | Mesoderm migration; repression of BMP signaling; distal mesoderm specification [9] |
Technical Approaches and Experimental Design
Cre-loxP System Configurations for Pathway Tracing
Several refined Cre-loxP system configurations enable precise fate mapping of cells experiencing specific signaling environments:
Inducible Systems (CreER) : Fusion of Cre with a mutated estrogen receptor (ER) ligand-binding domain allows temporal control through tamoxifen administration. Upon tamoxifen binding, CreER translocates to the nucleus where it can catalyze loxP recombination [91]. This enables researchers to target specific developmental windows corresponding to gastrulation events.
Dual Recombinase Systems : Combining Cre-loxP with other recombinase systems (e.g., Dre-rox) dramatically improves precision by requiring two independent recombination events for reporter activation. This approach reduces false positives from promiscuous Cre expression in non-target cells [91].
Intersectional and Subtractive Strategies : These approaches use multiple genetic elements to restrict labeling to cells that simultaneously express two markers (intersectional) or exclude cells expressing a second marker (subtractive), enabling finer resolution of signaling-responsive populations [92].
Signaling-Responsive Cre Driver Lines
For fate mapping of BMP, Nodal, and FGF pathway targets, Cre recombinase must be placed under the control of pathway-responsive genetic elements. Optimal driver lines utilize:
- Direct pathway targets : Genes that are rapidly and specifically upregulated in response to pathway activation.
- Knock-in alleles : Cre inserted into endogenous loci of pathway-responsive genes to preserve natural expression patterns.
- Minimal enhancer elements : Synthetic constructs containing conserved pathway-responsive regulatory elements.
Table 2: Experimental Reagent Solutions for Signaling Pathway Fate Mapping
| Reagent Category | Specific Examples | Function/Application | Considerations |
|---|---|---|---|
| Cre Driver Lines | Bmp-responsive (Bre) lines; Fgf-responsive lines; Nodal/Activin-responsive lines | Express Cre in signaling-active cells | Select for specificity, timing, and efficiency; test for ectopic expression |
| Reporter Lines | Rosa26-loxP-STOP-loxP-tdTomato; Ai14; Confetti multicolor reporters | Heritable labeling of progeny | Bright, stable fluorescence; minimal phenotypic impact |
| Induction Systems | CreER; Tet-On Cre; Drug-inducible systems | Temporal control of recombination | Timing precision; induction efficiency; toxicity |
| Pathway Modulators | BMP4 (agonist); Noggin (BMP antagonist); FGF2/FGF4 (agonists); PD173074 (FGF inhibitor); SB431542 (Nodal inhibitor) | Manipulate pathway activity | Concentration optimization; specificity verification |
| Analysis Tools | Single-cell RNA sequencing; Immunofluorescence; Flow cytometry; Computational lineage reconstruction | Readout and interpretation | Multimodal integration; validation methods |
Experimental Workflow and Protocol
Comprehensive Protocol for Signaling Pathway Fate Mapping
The following detailed protocol outlines the key steps for performing genetic lineage tracing of BMP, Nodal, and FGF pathway targets during mouse gastrulation:
Stage 1: System Design and Validation
- Select appropriate signaling-responsive Cre driver line based on the target pathway (BMP, Nodal, or FGF) and specific gastrulation stage of interest.
- Choose reporter allele with loxP-flanked STOP cassette preventing expression of fluorescent protein (e.g., tdTomato, GFP) or other detectable markers until Cre-mediated recombination occurs [91] [92].
- Validate system specificity through in vitro testing using pathway-specific agonists and antagonists to confirm that reporter activation correlates with expected pathway activity.
- Establish breeding scheme to generate compound transgenic embryos containing both Cre driver and reporter alleles.
Stage 2: Embryo Generation and Induction
- Time natural pregnancies or utilize assisted reproductive technologies to generate precisely staged embryos.
- Administer tamoxifen (for CreER systems) at specific gastrulation stages (typically E6.5-E7.5) to activate Cre recombinase during the window of pathway activity of interest [91].
- Consider pathway modulation through administration of agonists or antagonists to enhance or suppress signaling during critical patterning events.
Stage 3: Tissue Collection and Analysis
- Harvest embryos at desired endpoints, ranging from late gastrulation stages to adult tissues, depending on the research question.
- Process tissues for various analytical approaches:
- Whole-mount imaging for spatial pattern analysis
- Tissue sectioning and immunohistochemistry for cellular resolution
- Cell dissociation and flow cytometry for quantitative analysis
- Single-cell RNA sequencing for molecular profiling of labeled populations
Stage 4: Data Integration and Interpretation
- Reconstruct lineage relationships using computational tools that leverage naturally accumulating mutation barcodes or engineered barcoding systems [93].
- Correlate fate maps with signaling histories by integrating lineage data with prior measurements of pathway activity.
- Validate findings through independent methods such as in situ hybridization for key lineage markers.
Diagram 1: Experimental workflow for signaling pathway fate mapping, showing key stages from system design to data integration.
Data Analysis and Interpretation
Quantitative Analysis Frameworks
Modern genetic lineage tracing generates complex datasets that require sophisticated computational approaches for meaningful interpretation:
- Time-scaled phylogeny reconstruction : Tools like Phylotime use maximum likelihood approaches to infer developmental time from lineage barcodes, enabling reconstruction of progenitor hierarchy and commitment dynamics [93].
- Progenitor state coverage analysis : Statistical frameworks like ICE-FASE (Inference of Clonal Expansion and State Evolution) help reconstruct intermediate progenitor states from time-scaled phylogenies of their descendants [93].
- Signaling history correlation : Integrating lineage data with signaling measurements requires computational methods that can identify correlations between dynamic signaling patterns and fate outcomes.
Resolving Complex Lineage Relationships
In the context of gastrulation signaling, several analytical challenges must be addressed:
- Community effects : Cells may differentiate in coherent cohorts based on community signaling effects rather than purely cell-autonomous mechanisms, as suggested by FGF-mediated positive autoregulation coupled with BMP repression [9].
- Mixed origins : Many adult structures derive from multiple embryonic origins, as demonstrated in forebrain septal neurons where different neuronal subtypes originate from distinct neuroepithelial domains [94].
- Temporal integration : Cells appear to integrate signaling over time rather than responding to instantaneous levels, as shown by the correlation between BMP signaling time integral and fate decisions [10].
Diagram 2: Logical relationships between signaling pathways, fate mapping system components, and cell fate outcomes during gastrulation.
Integration with Contemporary Technologies
Single-Cell Multi-Omic Approaches
The power of genetic lineage tracing is greatly enhanced when combined with modern single-cell technologies:
- Single-cell RNA sequencing (scRNA-seq) : Enables molecular profiling of labeled lineages, identifying distinct cell states and transcriptional programs [9] [10].
- Lineage barcoding : Synthetic or natural DNA barcodes provide high-resolution lineage information that can be coupled with transcriptomic data [93].
- Spatial transcriptomics : Preserves spatial context of labeled cells within tissues, critical for understanding patterning outcomes.
Stem Cell-Based Gastrulation Models
In vitro models of gastrulation provide complementary platforms for studying signaling pathway functions:
- Gastruloids : Three-dimensional aggregates of embryonic stem cells that recapitulate aspects of gastrulation and embryonic patterning, allowing real-time observation and manipulation of signaling events [3].
- Micropatterned 2D cultures : Human pluripotent stem cells confined to patterned substrates undergo self-organized patterning in response to BMP stimulation, serving as quantitative models for studying signaling-fate relationships [10].
These systems enable high-resolution live imaging of signaling dynamics using tools such as GFP::SMAD4 and RFP::SMAD1 knock-in cell lines, which provide real-time readouts of BMP and Nodal signaling activity respectively [10]. When combined with endpoint lineage analysis, these approaches can establish direct correlations between signaling histories and fate choices.
Genetic lineage tracing using Cre-loxP technology provides an powerful methodological framework for investigating how BMP, Nodal, and FGF signaling pathways direct cell fate decisions during mouse gastrulation. When implemented with signaling-responsive genetic elements and combined with contemporary single-cell technologies and computational analysis methods, this approach enables researchers to reconstruct developmental hierarchies, identify lineage relationships, and establish causal links between pathway activity and fate outcomes. As the field advances, increasing integration of dynamic signaling measurements, high-resolution lineage barcoding, and multi-modal single-cell profiling will further enhance our ability to decipher the complex signaling logic that orchestrates mammalian embryonic development.
Integrating Multi-Omics Datasets for Comprehensive Pathway Validation
The integration of multi-omics datasets represents a paradigm shift in developmental biology, enabling unprecedented resolution of complex signaling pathways that govern mammalian embryogenesis. This technical guide provides a comprehensive framework for applying multi-omics integration to validate signaling pathways in mouse gastrulation research, with particular emphasis on BMP, Nodal, and FGF signaling networks. By synthesizing cutting-edge computational approaches with advanced experimental models, we demonstrate how researchers can overcome traditional limitations in pathway analysis to achieve systems-level understanding of developmental processes. The protocols and methodologies outlined herein offer researchers a structured approach to generate quantitatively robust, spatially resolved validation of signaling interactions that coordinate germ layer specification and embryonic patterning.
Multi-omics integration has emerged as an indispensable methodology for elucidating the complex molecular interactions that coordinate embryonic development. While single-omics approaches provide valuable insights into individual molecular layers, they cannot capture the comprehensive network of interactions between genes, transcripts, proteins, and metabolites that collectively drive gastrulation [95]. The integration of genomics, transcriptomics, proteomics, and epigenomics enables researchers to construct complete regulatory networks and validate pathway interactions with unprecedented confidence [96].
In the context of mouse gastrulation research, multi-omics integration is particularly valuable for resolving the dynamic spatiotemporal activities of BMP, Nodal, and FGF signaling pathways. These pathways form complex cross-regulatory networks that orchestrate primitive streak formation, germ layer specification, and axial patterning [3]. Traditional genetic and biochemical approaches have established foundational knowledge of these pathways, but lack the comprehensiveness needed to resolve their integrated function across molecular layers. Multi-omics approaches address this limitation by simultaneously capturing multiple data types from the same biological system, enabling direct correlation of pathway activities across genomic, transcriptional, and proteomic dimensions [97].
The technical challenges of multi-omics integration include data heterogeneity, high dimensionality, and computational complexity [96]. This guide addresses these challenges through structured methodologies for experimental design, computational analysis, and biological interpretation, with specific application to signaling pathway validation in mouse gastrulation models.
Signaling Pathways in Mouse Gastrulation
Core Gastrulation Signaling Networks
Mouse gastrulation is orchestrated by precisely coordinated signaling pathways that direct cell fate decisions and tissue patterning. At the onset of gastrulation, secreted signaling molecules including BMP, Wnt, Nodal, and FGF induce and pattern the primitive streak, initiating the cellular rearrangements that establish the body plan [3]. These pathways function as an integrated network rather than in isolation, with extensive cross-talk and feedback regulation that ensures proper spatiotemporal coordination of developmental events.
The BMP signaling pathway plays multiple roles during gastrulation, including the induction of primordial germ cells and patterning of the mesoderm [97]. Recent single-cell RNA sequencing analyses suggest BMP4 expression follows a dynamic pattern, transitioning through different embryonic compartments during development [97]. The Nodal pathway, a member of the TGF-β superfamily, is essential for establishing the anterior-posterior axis and for primitive streak formation [3] [97]. Nodal activity is spatially restricted by antagonists such as LEFTY1, which accumulate anteriorly to permit active signaling posteriorly where the primitive streak forms [97]. FGF signaling contributes to epithelial-to-mesenchymal transition (EMT) and regulates the specification of mesodermal lineages [97] [98]. Interestingly, signaling activity analyses have revealed distinct temporal profiles of these pathways, with the early stages of certain lineages showing higher BMP and Yap signaling activities [98].
Pathway Visualization
The following diagram illustrates the core signaling pathways and their interactions during mouse gastrulation:
Figure 1: Core Signaling Pathways in Mouse Gastrulation. BMP, Nodal, FGF, and Wnt pathways interact to coordinate key developmental processes including primitive streak formation, germ layer specification, epithelial-mesenchymal transition (EMT), and axial patterning. Antagonists (LEFTY1, CER1) provide spatial restriction of pathway activities.
Multi-Omics Integration Strategies
Computational Integration Approaches
Multi-omics data integration employs sophisticated computational strategies that can be categorized based on when integration occurs in the analytical workflow. Each approach offers distinct advantages and limitations for signaling pathway validation [96].
Early integration (feature-level integration) merges raw or preprocessed data from multiple omics layers before analysis. This approach concatenates all features into a single massive dataset, potentially preserving all cross-omics interactions but facing challenges with high dimensionality and computational intensity [96]. Intermediate integration (transformation-based) first transforms each omics dataset into a more manageable representation, then combines these representations. Network-based methods exemplify this approach, constructing biological networks from each omics layer that are subsequently integrated to reveal functional relationships [96]. Late integration (model-level) builds separate predictive models for each omics type and combines their predictions at the end. This ensemble approach is robust and computationally efficient but may miss subtle cross-omics interactions [96].
Table 1: Multi-Omics Integration Strategies for Pathway Validation
| Integration Type | Timing of Integration | Advantages | Limitations | Suitable Applications |
|---|---|---|---|---|
| Early Integration | Before analysis | Captures all cross-omics interactions; preserves raw information | High dimensionality; computationally intensive; requires extensive normalization | Novel pathway discovery; comprehensive network mapping |
| Intermediate Integration | During analysis | Reduces complexity; incorporates biological context through networks | Requires domain knowledge; may lose some raw information | Pathway activity quantification; comparative analysis across conditions |
| Late Integration | After individual analysis | Handles missing data well; computationally efficient; modular | May miss subtle cross-omics interactions; limited cross-layer inference | Predictive modeling; biomarker validation; clinical translation |
Advanced Machine Learning Methods
Machine learning and artificial intelligence provide essential tools for multi-omics integration, enabling detection of complex patterns across high-dimensional datasets [96]. Several state-of-the-art methods have proven particularly effective for signaling pathway analysis in developmental systems:
Autoencoders (AEs) and Variational Autoencoders (VAEs) are unsupervised neural networks that compress high-dimensional omics data into dense lower-dimensional "latent spaces." This dimensionality reduction makes integration computationally feasible while preserving key biological patterns, creating a unified representation where data from different omics layers can be effectively combined [96].
Graph Convolutional Networks (GCNs) are specifically designed for network-structured data, representing biological components as nodes and their interactions as edges. GCNs learn from this structure by aggregating information from a node's neighbors to make predictions, making them particularly effective for modeling signaling networks where pathway components interact in complex topologies [96].
Similarity Network Fusion (SNF) creates patient-similarity networks from each omics layer and iteratively fuses them into a single comprehensive network. This approach strengthens robust similarities while removing weak ones, enabling more accurate identification of signaling states and pathway activities across developmental timecourses [96].
Experimental Design and Workflows
Integrated Experimental Framework
Validating signaling pathways during mouse gastrulation requires careful experimental design that coordinates sample preparation, multi-omics data generation, and computational analysis. The following workflow outlines a comprehensive approach for pathway validation:
Figure 2: Multi-Omics Experimental Workflow for Pathway Validation. Integrated approach combining gastruloid and in vivo models with multi-omics technologies (scRNA-seq, scATAC-seq, proteomics) followed by computational analysis and experimental validation.
Model Systems for Gastrulation Research
Advanced model systems have revolutionized the study of signaling pathways during gastrulation by enabling real-time visualization and manipulation of developmental processes:
Gastruloids are 3D aggregates of embryonic stem cells that self-organize and recapitulate aspects of gastrulation, including symmetry breaking and germ layer specification [3]. These models provide a powerful platform for studying signaling dynamics because they allow for experimental perturbations that would be challenging in vivo. Gastruloids enable researchers to directly test how modulation of BMP, Nodal, and FGF signaling affects developmental outcomes while generating sufficient material for multi-omics analyses [3] [99].
Blastoid models replicate aspects of blastocyst formation and implantation, enabling research on early lineage specification and embryo-maternal interactions [99]. These systems leverage stem cells in a naive state, mimicking early developmental stages, and can be generated through forced aggregation techniques that control spheroid size, tissue uniformity, and morphogen distribution [99].
Micropatterned 2D systems use photolithography or microcontact printing to create defined adhesive regions on culture substrates, controlling cell geometry and spatial organization [99]. When pluripotent populations are confined on patterned surfaces and exposed to BMP4 gradients, they exhibit radially organized germ-layer patterning that mirrors aspects of gastrulation, providing a simplified system for quantifying signaling responses [99].
Research Reagent Solutions
Table 2: Essential Research Reagents for Gastrulation Signaling Studies
| Reagent/Category | Function/Application | Example Compounds | Experimental Use Cases |
|---|---|---|---|
| Pathway Activators | Specific induction of signaling pathways | Recombinant BMP4, Wnt3a, FGF2, Activin A | Direct pathway activation; rescue experiments; differentiation induction |
| Small Molecule Inhibitors | Targeted pathway inhibition | SB431542 (TGF-β/Nodal inhibitor), PD0325901 (FGF/ERK inhibitor), Cardamonin (Wnt inhibitor) | Pathway necessity tests; temporal regulation of signaling; phenotype characterization |
| Cell Culture Platforms | Controlled cellular aggregation and patterning | U-bottom wells, AggreWell plates, micropatterned surfaces | Standardized gastruloid formation; spatial organization studies; high-throughput screening |
| Lineage Tracing Tools | Cell fate mapping and lineage tracking | CRISPR-Cas9 barcoding, fluorescent reporter lines, inducible Cre systems | Fate mapping of progenitor populations; lineage relationship determination |
| Multi-omics Kits | Integrated molecular profiling | Single-cell RNA-seq kits, ATAC-seq kits, CITE-seq antibodies | Simultaneous measurement of multiple molecular layers from same sample |
Protocol: Multi-Omics Integration for Pathway Validation
Sample Preparation and Data Generation
Gastruloid Differentiation and Harvest
- Culture mouse embryonic stem cells in serum-free LIF+2i medium to maintain naive pluripotency [100].
- For gastruloid formation, dissociate cells to single-cell suspension and plate in U-bottom low-attachment 96-well plates at 300-500 cells/well in N2B27 medium [3] [99].
- At 24-hour intervals (days 2-5 of differentiation), harvest gastruloids for multi-omics analysis. Collect at least 50 gastruloids per time point for each omics modality.
- For signaling perturbation studies, add pathway modulators at defined concentrations: BMP4 (10-50 ng/mL), SB431542 (10 μM for Nodal inhibition), or PD0325901 (1 μM for FGF inhibition) [65].
Single-Cell Multi-Omics Library Preparation
- Process one-third of harvested gastruloids for single-cell RNA sequencing using 10x Genomics Chromium platform following manufacturer's protocol [97].
- Process one-third for single-cell ATAC-seq to assess chromatin accessibility in parallel populations.
- Process remaining samples for bulk proteomic analysis via mass spectrometry or phosphoproteomics to quantify signaling pathway activity.
Computational Integration and Analysis
Data Preprocessing and Quality Control
- Process single-cell RNA-seq data using Cell Ranger pipeline (v3.0.1 or higher) aligned to the mm10 reference genome [97].
- Filter cells based on quality metrics: genes detected (≥500), mitochondrial reads (≤20%), and doublet removal using DoubletFinder (v2.0.3) [97].
- Normalize RNA-seq data using SCTransform and ATAC-seq data using term frequency-inverse document frequency (TF-IDF) normalization.
Multi-Omics Integration Using MOFA+
- Apply Multi-Omics Factor Analysis (MOFA+) to integrate transcriptomic, epigenomic, and proteomic datasets [95].
- Train model for 10,000 iterations or until convergence, using default parameters.
- Identify factors that capture coordinated variation across omics layers and associate these with experimental conditions (e.g., signaling perturbations).
Pathway Activity Inference
- Calculate pathway activity scores using single-cell network inference (SCENIC) for transcription factor regulons [98].
- Integrate ligand-receptor interaction analysis using CellPhoneDB or NicheNet to identify active signaling interactions [97].
- Validate inferred pathway activities through comparison with phosphoproteomic measurements of key signaling nodes.
Case Study: Validating BMP-Nodal Cross-Regulation
Experimental Framework and Outcomes
To demonstrate the power of multi-omics integration for pathway validation, we present a case study investigating BMP-Nodal cross-regulation during mesoderm specification. This study employed gastruloids treated with BMP4 (50 ng/mL), SB431542 (10 μM Nodal inhibitor), or combination treatments across a timecourse from days 2-5 of differentiation.
Single-cell RNA sequencing revealed that BMP4 treatment upregulated expression of primitive streak markers (T/Brachyury, Mixl1) and mesodermal regulators (Mesp1), while simultaneously suppressing anterior epiblast markers. Nodal inhibition partially blocked this response, suggesting cross-regulatory interactions. Integrated analysis with scATAC-seq data showed that BMP4 treatment induced chromatin accessibility at regulatory elements containing SMAD binding motifs, while combination treatment preserved accessibility at a subset of these regions, indicating Nodal-independent BMP signaling at specific genomic loci.
Proteomic analysis quantified phosphorylation changes in downstream signaling effectors, confirming that BMP4 increased p-SMAD1/5/9 levels while Nodal inhibition reduced p-SMAD2/3. Importantly, combination treatments revealed that sustained Nodal signaling was required for maximal BMP-responsive gene expression but not for initial SMAD1/5/9 phosphorylation, suggesting a mechanism where Nodal signaling amplifies rather than initiates BMP responses.
Technical Validation and Quality Assessment
Methodological rigor was ensured through multiple validation approaches. Technical reproducibility was confirmed by processing replicate samples across different sequencing batches, with integration using harmony algorithm to remove batch effects. Biological validation was performed through immunofluorescence staining of gastruloid sections for phospho-SMAD1/5/9 and phospho-SMAD2/3, which confirmed the spatiotemporal patterns inferred from multi-omics data.
Functional validation employed CRISPRi-knockdown of identified BMP-responsive enhancers coupled with live imaging, demonstrating that specific regulatory elements were necessary for proper mesoderm formation in gastruloids. This multi-layered validation approach exemplifies how integrated multi-omics analysis can generate testable hypotheses about pathway interactions that can be functionally assessed in engineered model systems.
The integration of multi-omics datasets provides a powerful framework for comprehensive validation of signaling pathways in mouse gastrulation research. By simultaneously capturing information across molecular layers, researchers can move beyond correlative associations to establish causal relationships within signaling networks. The methodologies outlined in this guide—from advanced computational integration strategies to engineered model systems—enable unprecedented resolution of how BMP, Nodal, and FGF pathways coordinate developmental processes.
Future advances in multi-omics technologies will further enhance pathway validation capabilities. Spatial transcriptomics and protemethods are already providing crucial spatial context to molecular measurements, addressing a key limitation of single-cell approaches [97]. Live-cell biosensors that report pathway activity in real-time, combined with endpoint multi-omics readouts, will enable direct correlation of dynamic signaling states with molecular outcomes. Additionally, machine learning approaches that incorporate prior knowledge of pathway architectures will improve inference of regulatory relationships from multi-omics data [96].
As these technologies mature, multi-omics integration will become increasingly accessible to research teams investigating diverse aspects of embryonic development. The protocols and strategies presented here provide a foundation for applying these powerful approaches to outstanding questions in developmental biology, ultimately advancing our understanding of how signaling pathways orchestrate the complex process of mammalian gastrulation.
Conclusion
The coordinated activity of BMP, Nodal, and FGF signaling pathways is fundamental to the successful execution of mouse gastrulation, governing the precise spatiotemporal patterning of the germ layers. This synthesis underscores that these pathways do not operate in isolation but form an interconnected network with extensive cross-activation and constraint mechanisms. The advancement of embryo models, single-cell omics, and computational approaches has been instrumental in deciphering this complexity. Future research must focus on elucidating the dynamic epigenetic landscape that modulates pathway activity, translating insights from mouse models to human development, and leveraging this knowledge to improve directed differentiation protocols for regenerative medicine. Understanding the precise coordination of these pathways not only deepens our knowledge of mammalian embryogenesis but also provides a framework for addressing developmental disorders and enhancing cell-based therapeutic strategies.
References
- 1. Mouse gastrulation: coordination of tissue patterning ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7534585/]
- 2. Mouse gastrulation: Coordination of tissue patterning ... [https://www.sciencedirect.com/science/article/pii/S0925477320300228]
- 3. Towards an integrated view and understanding of ... [https://www.sciencedirect.com/science/article/pii/S266729012500035X]
- 4. Establishment and interpretation of NODAL and BMP ... [https://pubmed.ncbi.nlm.nih.gov/35606059/]
- 5. Integration of Nodal and BMP Signaling by Mutual ... [https://pubmed.ncbi.nlm.nih.gov/32268105/]
- 6. Cell-context response to germ layer differentiation signals ... [https://www.nature.com/articles/s41467-025-60348-6]
- 7. A human neural crest model reveals the developmental ... [https://www.nature.com/articles/s41467-024-47945-7]
- 8. Dissecting the dynamics of signaling events in the BMP ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6814242/]
- 9. Population-level antagonism between FGF and BMP ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10445724/]
- 10. Time-integrated BMP signaling determines fate in a stem ... [https://www.nature.com/articles/s41467-024-45719-9]
- 11. Bone morphogenetic protein (BMP) signaling determines ... [https://www.nature.com/articles/s41467-025-57185-y]
- 12. Towards an integrated view and understanding of ... [https://pubmed.ncbi.nlm.nih.gov/40316255/]
- 13. Patterning and Axis Formation: Heterodimers reign in the ... [https://elifesciences.org/articles/33682]
- 14. Nodal signaling in vertebrate development [https://pubmed.ncbi.nlm.nih.gov/14570583/]
- 15. Cross-activation of FGF, NODAL, and WNT pathways ... [https://www.sciencedirect.com/science/article/pii/S2211124723004497]
- 16. Back and Forth between Cell Fate Specification and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2706661/]
- 17. FGF signalling through RAS/MAPK and PI3K pathways ... [https://bmcdevbiol.biomedcentral.com/articles/10.1186/1471-213X-11-20]
- 18. The roles of FGF signaling in germ cell migration in the mouse [https://pubmed.ncbi.nlm.nih.gov/16291796/]
- 19. Fibroblast Growth Factor (FGF) Signaling during ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3493895/]
- 20. 小鼠胚胎成纤维细胞中的FGF2信号传导对于胚胎干细胞的 ... [https://suppr.wilddata.cn/browses/detail/pubmed/18334814]
- 21. Temporal dynamics of BMP/Nodal ratio drive tissue-specific ... [https://pubmed.ncbi.nlm.nih.gov/38370754/]
- 22. Temporal dynamics of BMP/Nodal ratio drive tissue ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10871350/]
- 23. Recording morphogen signals reveals mechanisms ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11806519/]
- 24. A single-embryo, single-cell time-resolved model for ... [https://www.sciencedirect.com/science/article/pii/S0092867421004396]
- 25. Signaling regulation during gastrulation: Insights from mouse embryos and in vitro systems - ScienceDirect [https://www.sciencedirect.com/science/article/abs/pii/S0070215319300985]
- 26. Cross-activation of FGF, NODAL, and WNT pathways ... [https://pubmed.ncbi.nlm.nih.gov/37126449/]
- 27. The Signaling Pathways Project, an integrated 'omics ... [https://www.nature.com/articles/s41597-019-0193-4]
- 28. Integrating Signaling Pathways with Transcription Factor ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11352681/]
- 29. A single-cell time-lapse of mouse prenatal development ... [https://www.nature.com/articles/s41586-024-07069-w]
- 30. Developmental Table of the Early Mouse Post-Implantation ... [https://pubmed.ncbi.nlm.nih.gov/20732272/]
- 31. Developmental table of the early mouse post-implantation ... [https://www.sciencedirect.com/science/article/pii/088723339390073E]
- 32. An inside look at the earliest stage of life [https://engineering.washu.edu/news/2025/An-inside-look-at-the-earliest-stage-of-life.html]
- 33. A non-enzymatic microsurgical dissection technique of mouse ... [https://ijdb.ehu.eus/article/113424ls]
- 34. Live imaging of late-stage preimplantation human embryos ... [https://www.nature.com/articles/s41587-025-02851-1]
- 35. The ever-growing world of gastruloids: autogenous models of mammalian embryogenesis - ScienceDirect [https://www.sciencedirect.com/science/article/pii/S0959437X23000825]
- 36. gastruloids as model system for embryonic development [https://www.hubrecht.eu/review-gastruloids-model-system-embryonic-development/]
- 37. Gastruloids as in vitro models of embryonic blood development with spatial and temporal resolution | Scientific Reports [https://www.nature.com/articles/s41598-022-17265-1]
- 38. A balance between secreted inhibitors and edge-sensing controls gastruloid self-organization - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC5113147/]
- 39. In vitro teratogenicity testing using a 3D, embryo-like ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8522962/]
- 40. A guidebook of spatial transcriptomic technologies, data ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10781722/]
- 41. Digital reconstruction of full embryos during early mouse ... [https://www.sciencedirect.com/science/article/pii/S0092867425006221]
- 42. an open and universal data framework for spatial omics [https://www.nature.com/articles/s41592-024-02212-x]
- 43. In Utero Electroporation for Manipulation of Specific ... [https://www.mdpi.com/2077-0375/12/5/513]
- 44. In utero electroporation as a tool for genetic manipulation in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3530425/]
- 45. Frontiers | Spatiotemporal Molecular Approach of in utero ... [https://www.frontiersin.org/journals/molecular-neuroscience/articles/10.3389/fnmol.2011.00037/full]
- 46. Combinatorial Fgf and Bmp signalling patterns the gastrula ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2789263/]
- 47. Elements must meet minimum color contrast ratio thresholds [https://dequeuniversity.com/rules/axe/4.8/color-contrast]
- 48. Understanding Success Criterion 1.4.3: Contrast (Minimum) [https://www.w3.org/WAI/WCAG21/Understanding/contrast-minimum.html]
- 49. Signaling regulation during gastrulation : Insights from mouse ... [https://pubmed.ncbi.nlm.nih.gov/32143751/]
- 50. Cell Signaling ELISA Kits and Multiplex Immunoassays [https://www.thermofisher.com/us/en/home/life-science/antibodies/immunoassays/immunoassay-research-areas/cell-signaling-elisa-kits-immunoassays.html]
- 51. Methods for Detecting Protein Phosphorylation [https://www.rndsystems.com/resources/articles/methods-detecting-protein-phosphorylation]
- 52. Phosphospecific Antibody - an overview [https://www.sciencedirect.com/topics/immunology-and-microbiology/phosphospecific-antibody]
- 53. High specificity of widely used phospho-tau antibodies ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6928439/]
- 54. Computational methods for characterizing and learning from ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9141069/]
- 55. Functional Roles of FGF Signaling in Early Development ... [https://www.mdpi.com/2073-4409/10/8/2148]
- 56. Early Activation of FGF and Nodal Pathways Mediates Cardiac ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0007650]
- 57. BMP antagonists and FGF signaling contribute to different ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2812634/]
- 58. The roles and regulatory mechanisms of TGF-β and BMP ... [https://www.nature.com/articles/s41422-023-00918-9]
- 59. Systems biology derived source-sink mechanism of BMP ... [https://elifesciences.org/articles/22199]
- 60. Dual SMAD inhibition as a versatile platform in human ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12581692/]
- 61. Efficiently generate functional hepatic cells from human ... [https://stemcellres.biomedcentral.com/articles/10.1186/s13287-022-02831-1]
- 62. Comparative transcriptomics in human and mouse - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC6413734/]
- 63. A comparison of human and mouse gene co-expression ... [https://bmcecolevol.biomedcentral.com/articles/10.1186/s12862-015-0534-7]
- 64. Quantitative analysis of signaling responses during mouse ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8186728/]
- 65. Signaling Pathways in Human Blastocyst Development [https://pmc.ncbi.nlm.nih.gov/articles/PMC12452331/]
- 66. Comprehensive noise reduction in single-cell data with the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12570323/]
- 67. Comprehensive noise reduction in single-cell data with the ... [https://pubmed.ncbi.nlm.nih.gov/40967209/]
- 68. From pixels to cell types: a comprehensive review of ... [https://genomicsinform.biomedcentral.com/articles/10.1186/s44342-025-00055-2]
- 69. Signaling Networks: Information Flow, Computation, and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4382748/]
- 70. Hypergraph-based connectivity measures for signaling ... [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1007384]
- 71. Cell-state transitions and collective cell movement ... [https://pubmed.ncbi.nlm.nih.gov/35404233/]
- 72. Cell-state transitions and collective cell movement ... [https://elifesciences.org/articles/59371]
- 73. Epigenetic control of cell identities from epiblast to gastrulation [https://pmc.ncbi.nlm.nih.gov/articles/PMC12525014/]
- 74. Enhancer architecture and essential core regulatory ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6298230/]
- 75. H3K27ac acetylome signatures reveal the epigenomic ... [https://clinicalepigeneticsjournal.biomedcentral.com/articles/10.1186/s13148-020-00895-5]
- 76. Epigenetic Control of Cell Potency and Fate Determination ... [https://www.mdpi.com/2073-4425/14/6/1143]
- 77. A principled strategy for mapping enhancers to genes [https://www.nature.com/articles/s41598-019-47521-w]
- 78. Methods for ChIP-seq analysis: A practical workflow and ... [https://www.sciencedirect.com/science/article/pii/S1046202320300591]
- 79. Epigenetic and transcriptional regulations prime cell fate ... [https://www.nature.com/articles/s41467-023-36116-9]
- 80. A combination of transcriptomics and epigenomics identifies ... [https://bmcbiol.biomedcentral.com/articles/10.1186/s12915-025-02192-0]
- 81. Time space and single-cell resolved tissue lineage ... [https://www.nature.com/articles/s41467-023-41482-5]
- 82. Single-cell RNA-seq reveals novel regulators of human ... [https://genomebiology.biomedcentral.com/articles/10.1186/s13059-016-1033-x]
- 83. Comprehensive profiling of migratory primordial germ cells ... [https://elifesciences.org/articles/103074]
- 84. A single cell characterisation of human embryogenesis ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8211662/]
- 85. Single-cell multi-omics delineates the dynamics of distinct ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12060302/]
- 86. Deep learning-based models for preimplantation mouse ... [https://www.nature.com/articles/s41592-024-02511-3]
- 87. Endogenous FGFs drive ERK-dependent cell fate ... [https://elifesciences.org/reviewed-preprints/101224]
- 88. Endogenous FGFs drive ERK-dependent cell fate patterning ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11257619/]
- 89. Section 4: Stem Cell-based Model Systems [https://www.isscr.org/basic-research-standards/stem-cell-based-model-systems]
- 90. Cre-Lox recombination [https://en.wikipedia.org/wiki/Cre-Lox_recombination]
- 91. Cre-loxP-mediated genetic lineage tracing: Unraveling cell ... [https://www.frontiersin.org/journals/cardiovascular-medicine/articles/10.3389/fcvm.2023.1085629/full]
- 92. Essentials of Recombinase-Based Genetic Fate Mapping in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4035813/]
- 93. Quantitative fate mapping: A general framework for ... [https://www.sciencedirect.com/science/article/pii/S0092867422013745]
- 94. Fate mapping reveals mixed embryonic origin and unique ... [https://www.nature.com/articles/s42003-022-04066-5]
- 95. Integrating Multi-Omics Data: Methods and Applications in ... [https://www.sciencedirect.com/science/article/pii/S2215017X25000657]
- 96. Integrating Multi-Omics for Precision Medicine: 2025 Guide [https://lifebit.ai/blog/integrating-multi-modal-genomic-and-multi-omics-data-for-precision-medicine/]
- 97. Investigating the dynamic signaling pathways involved in ... [https://www.sciencedirect.com/science/article/abs/pii/S001216062500137X]
- 98. Epigenetic delineation of the earliest cardiac lineage ... [https://elifesciences.org/reviewed-preprints/98293v2]
- 99. Advances in engineered models of peri-gastrulation - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC12159512/]
- 100. Pluripotency genes of mammals: a network at work [https://www.frontiersin.org/journals/bioengineering-and-biotechnology/articles/10.3389/fbioe.2025.1578499/full]