Primitive Streak EMT: From Gastrulation Mechanisms to Cancer Metastasis
This article provides a comprehensive analysis of the epithelial-mesenchymal transition (EMT) within the primitive streak, the foundational event of gastrulation.
Primitive Streak EMT: From Gastrulation Mechanisms to Cancer Metastasis
Abstract
This article provides a comprehensive analysis of the epithelial-mesenchymal transition (EMT) within the primitive streak, the foundational event of gastrulation. We explore the evolutionary-conserved signaling pathways and transcriptional regulators that drive this primary EMT to form the mesoderm and endoderm. For researchers and drug development professionals, the review connects these developmental mechanisms to pathological processes, particularly cancer metastasis and therapy resistance. It further details modern methodologies for studying EMT, discusses challenges in interpreting its plasticity, and evaluates emerging therapeutic strategies that target EMT pathways, offering a translational perspective from embryo to patient.
The Primitive Streak: Unraveling the Foundational EMT Event in Gastrulation
Epithelial-mesenchymal transition (EMT) within the primitive streak represents a fundamental biological process that drives gastrulation and the establishment of the three germ layers in amniote embryos. This whitepaper traces the evolution of EMT concepts from initial phenomenological observations to contemporary molecular definitions, synthesizing current understanding of the signaling networks, cellular mechanisms, and experimental methodologies that define this critical developmental event. Within the context of broader EMT research, the primitive streak offers a paradigm for studying how epithelial cells undergo controlled phenotypic changes to acquire migratory potential, a process whose reactivation in adulthood contributes to pathological conditions including cancer metastasis and fibrosis. This technical guide provides researchers with a comprehensive framework for investigating EMT within this specific developmental context, highlighting conserved molecular principles and established experimental approaches.
The primitive streak is a transient embryonic structure that forms in the early amniote embryo (including humans, mice, and chicks) and serves as the architectural foundation for gastrulation [1]. This linear structure emerges at the posterior midline of the bilaminar embryonic disc and establishes the cranial-caudal and left-right body axes [1]. Anatomically, the primitive streak extends through the midline, with its cranial end expanding to form the primitive node (Hensen's node in birds), which contains the primitive pit that extends caudally as the primitive groove [1].
Functionally, the primitive streak provides the portal through which epiblast cells ingress during gastrulation, undergoing EMT to form the mesoderm and definitive endoderm [2] [1]. The epiblast, a single epithelial layer of the bilaminar embryonic disc, serves as the source of all embryonic material in amniotes, with a subset of its cells giving rise to the primitive streak [1]. During this process, epithelial cells downregulate epithelial characteristics, including cell-cell junctions and apico-basal polarity, and acquire mesenchymal features that enable migration and invasion into the underlying extracellular matrix [3] [2].
The significance of EMT at the primitive streak extends beyond developmental biology, as the molecular programs governing this process are reactivated in pathological contexts. Cancer biologists have demonstrated that EMT contributes to malignancy-associated phenotypes, notably invasiveness, dissemination, and therapeutic resistance [3] [4]. Thus, understanding the precise mechanisms governing EMT within the primitive streak provides fundamental insights with broad implications for both developmental and disease processes.
Historical Evolution of EMT Concepts
Initial Observations and Terminology
The conceptual foundation for EMT emerged from pioneering embryological studies in the mid-20th century. Elizabeth Hay and colleagues provided the first definitive descriptions of this process during embryonic development, initially terming it "epithelial-mesenchymal transformation" [5]. Using chick embryo models and electron microscopy, Hay documented how epithelial cells transformed into mesenchymal cells during neural tube formation and migration of neural crest cells [5]. These early observations emphasized the dramatic changes in cellular architecture and behavior, focusing on morphological transitions rather than molecular mechanisms.
The terminology evolved from "transformation" to "transition" to distinguish the process from neoplastic transformation and to reflect its reversible nature, particularly after the identification of mesenchymal-epithelial transition (MET) during development [3] [5]. This semantic shift followed the first meeting of the EMT International Association (TEMTIA) in 2003, which standardized nomenclature across the field [3].
From Morphological Descriptions to Molecular Mechanisms
Early research relied predominantly on microscopic observations of cellular shape changes. Hay's seminal work with chick lens epithelial cells demonstrated that when suspended in collagen gels, these cells developed cytoplasmic extensions resembling pseudopods and acquired migratory capability, mirroring mesenchymal behavior [5]. Simultaneously, studies in mouse embryogenesis documented that the first mesenchymal cells appearing during development lost desmosomes and cytokeratin expression while gaining vimentin expression [5].
The molecular era of EMT research began in the 1990s with the identification of specific growth factors and transcription factors governing the process. Transforming growth factor beta (TGF-β) emerged as a potent inducer of EMT across multiple systems [3] [5]. Landmark studies in Drosophila developmental genetics identified Snail and Twist as master regulators of mesoderm formation during gastrulation, with subsequent work revealing the conservation of these transcription factors in chordates [3]. The discovery that Slug (Snai2) could convert epithelial carcinoma cells into mesenchymal derivatives established a direct connection between developmental EMT and cancer progression [3].
Modern Definitions and Consensus Framework
TEMTIA Consensus Guidelines
The exponential growth in EMT research—with over 5,700 publications in 2019 alone—highlighted the need for standardized definitions and guidelines [3]. In 2020, the EMT International Association (TEMTIA) published a consensus statement establishing a unified framework for EMT research. This consensus defined EMT as "a cellular process during which epithelial cells acquire mesenchymal phenotypes and behaviour following the downregulation of epithelial features" [3].
The guidelines emphasize that EMT encompasses dynamic changes in cellular organization rather than a simple binary switch. The epithelial state is characterized by stable cell-cell junctions, apical-basal polarity, and interactions with the basement membrane, while the mesenchymal state features fibroblast-like morphology, cytoskeletal reorganization, and increased migratory capacity [3].
The Spectrum of EMT States
A critical advancement in modern EMT understanding is the recognition of phenotypic plasticity and the existence of multiple intermediate states along the epithelial-mesenchymal spectrum [3] [4]. The primitive streak exemplifies this plasticity, as cells undergoing EMT often reside in intermediate or "hybrid" states that simultaneously express both epithelial and mesenchymal characteristics [3].
Table 1: Characteristics of Cellular States During EMT in the Primitive Streak
| Cellular State | Epithelial Features | Mesenchymal Features | Functional Capabilities |
|---|---|---|---|
| Epithelial (Pre-ingression) | Stable junctions, apical-basal polarity, basement membrane attachment | None | Proliferation, epithelial barrier function |
| Hybrid/Intermediate | Residual junctions, partial polarity | Cytoskeletal reorganization, initiated motility | Apical constriction, beginning of delamination |
| Mesenchymal (Post-ingression) | No junctions, no polarity | Fibroblast-like morphology, front-rear polarity | Individual migration, ECM invasion |
This spectrum model explains the diversity of EMT manifestations observed in different biological contexts and has necessitated a move away from rigid, binary classifications [3]. For primitive streak EMT, this means recognizing that ingressing cells may display varying degrees of epithelial and mesenchymal characteristics throughout the process.
Molecular Regulation of EMT in the Primitive Streak
Signaling Pathways
The formation of the primitive streak and initiation of EMT are coordinated by a conserved network of signaling pathways that operate in a spatially and temporally regulated manner.
Table 2: Key Signaling Pathways in Primitive Streak Formation and EMT
| Signaling Pathway | Key Ligands/Components | Function in Primitive Streak EMT | Experimental Evidence |
|---|---|---|---|
| Wnt Signaling | Wnt3, Wnt8C, β-catenin | Confers competence for streak formation; regulates EMT transcription factors | Wnt3 deletion abolishes streak formation; ectopic Wnt induces multiple streaks [2] [1] |
| Nodal/TGF-β Signaling | Nodal, Vg1, TGF-β receptors | Induces primitive streak formation; activates EMT transcriptional programs | Nodal mutants fail to gastrulate; Vg1 misexpression induces ectopic streaks [2] [1] |
| FGF Signaling | FGF8, FGFR1 | Maintains EMT regulatory network; supports mesoderm migration | FGFR1-deficient mice form streaks but arrest mesoderm production [2] [1] |
| BMP Signaling | BMP4, BMP7, Chordin | Spatial restriction of streak formation; modulated by inhibitors | BMP misexpression prevents streak formation; Chordin induces ectopic streaks [1] |
Diagram 1: Signaling pathways regulating EMT in the primitive streak. Multiple extracellular signals converge on EMT transcription factors that orchestrate cellular changes required for ingression.
Transcriptional Control
EMT at the primitive streak is orchestrated by core transcription factors (EMT-TFs) that directly repress epithelial genes and activate mesenchymal programs. The Snail family (Snail and Slug/Snai2) plays particularly critical roles, with demonstrated functions in repressing E-cadherin expression and facilitating junction disassembly [3] [2]. Additional EMT-TFs including Twist, Zeb1, and Zeb2 cooperate to establish the mesenchymal phenotype [3]. These factors typically function in a network rather than individually, with extensive cross-regulation and contextual specificity.
In mouse embryos, Snail expression marks the definitive primitive streak domain and correlates spatiotemporally with basement membrane breakdown and cell ingression [6]. The transcriptional program activated by these factors leads to cytoskeletal reorganization, expression of matrix-degrading enzymes, and changes in cell-matrix adhesion molecules that collectively enable delamination and migration.
Epigenetic Regulation
Beyond transcriptional control, EMT involves complex epigenetic regulation that enables dynamic and reversible phenotypic changes. DNA methylation, histone modifications, non-coding RNAs, chromatin remodeling, and 3D genome architecture collectively orchestrate transcriptional plasticity across the EMT spectrum [4]. These regulatory layers allow cells to stabilize intermediate states or undergo complete transition to a mesenchymal phenotype in response to developmental cues.
Cellular Dynamics and Morphological Changes
Apical Constriction Mechanism
At the cellular level, EMT in the primitive streak involves a stereotypical sequence of morphological changes. High-resolution live imaging of mouse embryos has revealed that epiblast cells undergo apical constriction in a pulsed, ratchet-like fashion through asynchronous shrinkage of apical junctions [6]. This process occurs asynchronously, with individual cells or small groups ingressing in a scattered pattern throughout the primitive streak domain [6].
During a 1-hour observation window, approximately 44% of cells within the primitive streak domain constrict and ingress, with about 48% ingressing as isolated events and 52% as pairs or small groups [6]. This asynchronous behavior ensures continuous epithelial integrity while permitting sustained cell emigration from the epiblast layer.
Cytoskeletal and Junctional Reorganization
The ratchet-like apical constriction is mediated by reciprocal enrichment of actomyosin networks and polarity proteins, particularly Crumbs2, at apical junctions [6]. This anisotropic distribution creates mechanical forces that drive junctional shrinkage. Simultaneously, cells elongate basally and eventually detach from the epithelial layer following breakdown of the underlying basement membrane, a hallmark of the primitive streak region [3] [6].
Live imaging studies using ZO-1-GFP reporters (marking tight junctions) have demonstrated that ingressing cells maintain junctional connections until late stages of constriction, with complete dissolution occurring only immediately before or during basal emigration [6]. This careful regulation prevents premature loss of epithelial integrity while facilitating individual cell delamination.
Experimental Models and Methodologies
Embryo Model Systems
The study of EMT in the primitive streak has leveraged multiple model systems, each offering distinct advantages:
- Chick embryos: Provide accessibility for micromanipulation and live imaging; primitive streak formation occurs in a flat disc ideal for observation [2] [1]
- Mouse embryos: Offer genetic tractability and relevance to mammalian development; recent advances in ex utero culture enable live imaging of gastrulation stages [6]
- Stem cell-based models: Human pluripotent stem cells can form 2D micropatterned colonies or 3D embryoids that recapitulate aspects of primitive streak formation and EMT [7]
Each system presents limitations, including optical inaccessibility in intact mouse embryos and incomplete representation of tissue interactions in stem cell models. Nevertheless, complementary use of these systems has generated a comprehensive understanding of primitive streak EMT.
Key Experimental Approaches
Modern analysis of primitive streak EMT employs multidisciplinary approaches:
- Live imaging: Using fluorescent reporters for junctional proteins (ZO-1-GFP), membranes (Rosa26mT/mG), or cytoskeletal components to visualize dynamic cellular behaviors [6]
- Genetic perturbation: Conditional knockout models to assess function of specific genes in spatial and temporal context [6]
- Signaling modulation: Application of pathway-specific agonists or antagonists to dissect regulatory networks [2] [1]
- Transcriptomic analysis: Single-cell RNA sequencing to characterize intermediate states and heterogeneity within the primitive streak population
Diagram 2: Experimental workflow for studying EMT in the primitive streak. Integrated approaches combine model systems with perturbation strategies and readouts to dissect molecular and cellular mechanisms.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Studying Primitive Streak EMT
| Reagent Category | Specific Examples | Function/Application | Experimental Use |
|---|---|---|---|
| Lineage Tracing & Live Imaging | ZO-1-GFP fusion protein | Labels tight junctions; visualizes apical surface dynamics | Quantitative analysis of apical constriction dynamics [6] |
| Rosa26mT/mG reporter | Membrane-localized fluorescent reporter; marks entire plasma membrane | Identification of completed ingression events [6] | |
| Signaling Modulators | Recombinant BMP4 | Induces EMT and germ layer patterning in stem cell models | Formation of micropatterned colonies with primitive streak-like organization [7] |
| SU5402 (FGF receptor inhibitor) | Inhibits FGF signaling; blocks downstream EMT events | Testing FGF requirement in primitive streak formation [1] | |
| Genetic Tools | Crb2 mutants | Loss-of-function model for polarity protein | Demonstrates Crumbs2 requirement in apical constriction and myosin localization [6] |
| Snail/Snail mutants | Target core EMT transcription factors | Assess necessity for primitive streak formation and cell ingression [3] | |
| Cell Culture Models | hPSC micropatterned colonies | 2D model of gastrulation with self-organized patterning | Study of human EMT in controlled, reproducible system [7] |
| Mouse epiblast stem cells (EpiSCs) | Model post-implantation epiblast | Investigation of EMT regulation in vitro [8] |
Future Directions and Clinical Implications
The study of EMT in the primitive streak continues to evolve with emerging technologies and conceptual frameworks. Single-cell multi-omics approaches are revealing unprecedented resolution of cellular heterogeneity and transitional states during gastrulation. Advanced imaging techniques, including light-sheet microscopy of cleared embryos, provide spatial context for molecular changes.
From a translational perspective, understanding the core principles of developmental EMT informs strategies for targeting pathological EMT in disease contexts. The recognition that cells can occupy stable intermediate states rather than completing a full transition suggests therapeutic approaches might aim to modulate plasticity rather than completely inhibit EMT. Furthermore, the conservation of molecular regulators between developmental and pathological EMT provides validated targets for intervention.
Stem cell-based embryo models offer ethically accessible platforms for probing human-specific aspects of primitive streak EMT, though researchers must remain cognizant of their limitations compared to in vivo systems [7]. As these models become more sophisticated, they will increasingly enable investigation of human gastrulation and associated EMT events without the technical and ethical constraints of embryo research.
EMT at the primitive streak represents a foundational process in embryonic development that has shaped our understanding of cellular plasticity across biological contexts. From initial morphological descriptions to contemporary molecular definitions, the conceptualization of this process has evolved significantly, recognizing the dynamic spectrum of intermediate states rather than a simple binary switch. The coordinated action of signaling pathways, transcription factors, and epigenetic regulators orchestrates precise cellular changes that enable gastrulation while maintaining tissue integrity. Continued investigation using sophisticated model systems and experimental approaches will further elucidate the mechanisms governing this essential process, with broad implications for both developmental biology and disease pathogenesis.
Epithelial-mesenchymal transition (EMT) represents a fundamental evolutionary mechanism driving morphogenesis across the metazoan kingdom. This whitepaper examines the conserved core principles of EMT from diploblastic organisms through triploblastic amniotes, with particular emphasis on primitive streak formation. We synthesize current understanding of EMT signaling pathways, transcriptional regulators, and cellular dynamics, highlighting both deeply conserved mechanisms and lineage-specific adaptations. Quantitative analyses of EMT dynamics, detailed experimental methodologies, and comprehensive reagent solutions provide researchers with practical tools for investigating these evolutionarily ancient processes in modern experimental systems.
Epithelial-mesenchymal transition (EMT) encompasses dynamic changes in cellular organization from epithelial to mesenchymal phenotypes, leading to functional alterations in cell migration and invasion [9]. The term "mesenchyme" itself (Greek "mesos" [middle] + "enkhuma" [infusion]) was initially coined by Hertwig and Hertwig to designate the fluid-like structure between ectodermal and endodermal cells in the oldest metazoans, including diploblastic species such as jellyfish [4]. This evolutionary perspective reveals EMT as a fundamental, conserved mechanism underlying the morphogenesis of nearly all metazoans, with its core regulatory components maintained across vast evolutionary timescales.
The conservation of EMT programs extends beyond molecular machinery to encompass functional roles in establishing body plans. In all multicellular organisms, cell division in the fertilized egg leads to cell collectives that assemble as epithelial-like primordia using an evolutionarily conserved molecular toolkit [4]. Complex evolutionary-conserved morphogenetic movements in gastrulation exemplify the extent of epithelial cell plasticity in the animal kingdom, yet remarkably, a single-gene knockout can modify the mode of gastrulation while achieving the same body plan [4]. This highlights both the robustness and plasticity of EMT mechanisms throughout evolution.
EMT Conservation Across Evolutionary Lineages
EMT in Diploblastic Organisms
Diploblastic organisms such as cnidarians (jellyfish and sea anemones) provide crucial insights into the evolutionary origins of EMT. As one of the most ancient phyla, cnidarians undergo gastrulation through several distinct mechanisms, including involution and invagination [4]. In these early-branching metazoans, the process of gastrulation can occur via invagination of part of the primitive epithelium without forming individual mesenchymal cells, illustrating an ancient form of EMT that may represent the evolutionary precursor to more complex EMT programs in triploblasts [4].
The conservation of EMT regulatory programs in diploblasts is evidenced by molecular studies revealing that cnidarians possess homologs of key EMT-related transcription factors and signaling pathways found in bilaterians. These conserved genetic programs enable the fundamental remodeling of epithelial tissues during development, establishing the basic body plan through processes that would be elaborated upon in more derived lineages.
EMT in Triploblastic Amniotes
In triploblastic amniotes (reptiles, birds, and mammals), EMT undergoes significant elaboration while maintaining core conserved elements. Morphologically recognizable mesenchymal cells are observed after gastrulation in triploblasts, with the process becoming more complex and regulated [4]. The amniote primitive streak represents a key evolutionary innovation, providing a spatially organized site for the coordinated ingression of mesoderm and endoderm progenitors.
Table 1: Evolutionary Conservation of EMT Features Across Metazoans
| Feature | Diploblasts | Triploblasts/Amniotes | Conservation Status |
|---|---|---|---|
| Gastrulation Mechanism | Invagination, involution | Primitive streak formation, ingression | Partially conserved |
| Transcription Factors | Snail, Twist homologs | Snail, Twist, ZEB families | Highly conserved |
| Signaling Pathways | TGF-β, Wnt, FGF | TGF-β, Wnt, FGF, BMP | Highly conserved |
| Cell Adhesion Changes | Cadherin switching | E- to N-cadherin transition | Conserved mechanism |
| Basement Membrane | Local degradation | Regulated breakdown | Conserved process |
| Plasticity | High | Context-dependent | Conserved principle |
Comparative analyses across amniote species reveal both conserved and divergent EMT strategies. For instance, the mouse primitive streak forms in situ by initiation of EMT without migration of a cell population, unlike chick and rabbit models which display convergence and extension behavior [10]. This illustrates how core EMT programs can be deployed differently across lineages while achieving similar morphogenetic outcomes.
Cellular and Molecular Mechanisms of EMT in Amniote Gastrulation
Signaling Pathways Governing Primitive Streak Formation
The initiation of EMT during amniote gastrulation is coordinated by an evolutionarily conserved network of signaling pathways. In mouse and chick embryos, the convergence of Wnt, BMP, and Nodal signals, together with FGF signaling, triggers the EMT process at the primitive streak [6] [11]. Wnt signaling appears to render cells competent to respond to other extracellular signals that initiate EMT, as demonstrated by the failure of primitive streak formation in Wnt3-deficient mice [2]. The TGF-β superfamily proteins Nodal and Vg1 are key inducers of gastrulation across species, with ectopic Vg1 expression alone being sufficient to induce additional primitive streaks in chick blastulae [2].
Table 2: Quantitative Dynamics of EMT in Mouse Primitive Streak
| Parameter | Value | Measurement Context | Citation |
|---|---|---|---|
| Ingression Rate | 44 ± 2% of cells/hour | Within primitive streak domain | [6] |
| Isolated Ingression | 48% | Percentage of single-cell events | [6] |
| Collective Ingression | 52% | Pairs or groups (3-4 cells) | [6] |
| Spatial Domain | ~40 μm region | Posterior midline primitive streak | [6] |
| Basal Lamina Breakdown | First step | Correlated with ingression | [10] |
| Apical Constriction | Pulsed, ratchet-like | Preceding ingression | [6] |
BMP signaling represents another crucial conserved pathway in EMT regulation. Multiple Bmp genes (Bmp2/4/7) are expressed in the primitive streak along its anteroposterior axis, with BmprIa-null mutant mice unable to initiate gastrulation [12]. BMP signaling appears to regulate EMT partially through induction of Snail family gene expression, with SMAD binding sites identified in Snail/Slug promoter regions [12]. The conservation of this BMP-EMT regulatory module across diverse species underscores its fundamental importance in morphogenesis.
Transcription Factor Networks
Downstream of signaling events, a core set of transcription factors executes the EMT program. The Snail and Twist transcription factors, first identified in Drosophila melanogaster as essential regulators of mesoderm formation, are conserved across metazoans including sea urchins, chicks, and mice [4] [11]. In amniotes, Snail1 transcription is activated as cells ingress to form mesoderm, where it downregulates E-cadherin transcription [11]. The functional conservation of these factors is remarkable, with Snail family members capable of promoting EMT across diverse phylogenetic contexts.
The EMT transcriptional network extends beyond Snail and Twist to include ZEB1, ZEB2, and other factors that operate in context-dependent combinations. These transcription factors collectively repress epithelial genes such as those encoding E-cadherin, occludins, and claudins, while activating mesenchymal genes including N-cadherin and various matrix components [13] [2]. The deployment of this core transcriptional machinery in different combinatorial patterns across species and developmental contexts illustrates how conserved components can be reconfigured to generate phenotypic diversity.
Cellular Dynamics and Cytoskeletal Reorganization
Live imaging of EMT in mouse and chick embryos has revealed highly conserved cellular dynamics during gastrulation. In the mouse embryo, epiblast cells undergo apical constriction in a pulsed, ratchet-like fashion through asynchronous shrinkage of apical junctions [6]. This process involves anisotropic and reciprocal enrichment of members of the actomyosin network and Crumbs2 complexes, which regulate the asynchronous shrinkage of cell junctions [6].
The conservation of actomyosin-mediated apical constriction from Drosophila to mice, despite divergent timescales (~1 hour vs. >24 hours) and spatial parameters, highlights the deep evolutionary conservation of this biomechanical process [6] [11]. Across species, the contraction of apical actomyosin networks generates the forces necessary for cell shape changes and tissue remodeling during EMT, with specific modulations according to developmental context.
Figure 1: Conserved EMT Regulatory Network. Core signaling pathways and transcription factors regulating EMT across diploblasts to amniotes. The network architecture is conserved, though specific components may vary between species.
Experimental Models and Methodologies
Model Systems for EMT Research
The study of evolutionarily conserved EMT mechanisms employs diverse model systems, each offering unique advantages. Mouse embryos provide insights into mammalian gastrulation, with the primitive streak forming at approximately embryonic day 6.5 (E6.5) [6]. Chick embryos offer accessibility for manipulation and imaging, with primitive streak formation occurring during the first days of incubation [2]. Drosophila embryos remain invaluable for genetic dissection of core EMT mechanisms, particularly in mesoderm formation [11]. Complementary insights come from zebrafish, Xenopus, and even cnidarian models, which collectively enable comparative analyses of EMT program evolution.
Table 3: Essential Research Reagents for EMT Investigation
| Reagent Category | Specific Examples | Function/Application | Context of Use |
|---|---|---|---|
| Lineage Reporters | ZO-1-GFP, Rosa26mT/mG | Visualization of cell membranes and junctions | Live imaging of EMT dynamics [6] |
| Signaling Modulators | Recombinant BMP4, Noggin, FGF inhibitors | Pathway activation/inhibition | Functional tests of signaling requirements [12] |
| Transcription Factor Tools | Snail1/Snail2 mutants, Snail overexpression constructs | Manipulation of EMT master regulators | Assessing EMT requirement and sufficiency [11] |
| Cell Adhesion Markers | E-cadherin antibodies, N-cadherin reporters | Tracking adhesion molecule dynamics | Monitoring epithelial vs mesenchymal states [2] |
| Cytoskeletal Reporters | Lifeact-GFP, Myosin II-GFP | Visualization of actin and myosin dynamics | Analysis of cell shape changes [6] |
| Metabolic Probes | Glucose uptake sensors, mitochondrial dyes | Assessment of metabolic changes | Correlation of metabolism with EMT states [4] |
Live Imaging and Quantitative Analysis Protocols
Modern analysis of EMT dynamics relies heavily on live imaging approaches. For mouse embryo imaging, protocols typically involve explant culture systems that maintain embryo viability while enabling high-resolution microscopy [6]. A standard methodology includes:
Embryo Preparation: Dissect E6.5-E7.5 mouse embryos in pre-warmed DMEM/F-12 medium supplemented with serum replacement [6].
Reporter Expression: Utilize transgenic reporters such as ZO-1-GFP for junctional labeling or membrane-targeted fluorescent proteins for whole-cell visualization [6].
Imaging Setup: Mount embryos in glass-bottom dishes with appropriate culture media maintained at 37°C with 5% CO2 [6].
Time-Lapse Acquisition: Collect z-stacks at 3-10 minute intervals using spinning disk or two-photon microscopy to balance resolution and phototoxicity [6] [14].
Image Analysis: Employ segmentation software to track individual cells, quantify apical surface area dynamics, and measure ingression timing [6].
For quantitative analysis of EMT dynamics, key parameters include apical surface area reduction rate, ingression timing relative to neighbors, migration speed post-ingression, and correlation with molecular marker expression [6]. The scattered, apparently stochastic ingression of epiblast cells in the mouse primitive streak necessitates tracking large cell numbers for statistical power [6].
Figure 2: Experimental Workflow for EMT Analysis. Key steps in analyzing EMT dynamics in model organisms, integrating live imaging with molecular validation.
Discussion: Evolutionary Implications and Future Directions
The conservation of EMT mechanisms from diploblasts to amniotes underscores the fundamental nature of this process in metazoan evolution. The core regulatory modules involving Wnt, TGF-β, and FGF signaling, coupled with Snail/Twist transcription factors, appear to have been established early in animal evolution and maintained with modifications across lineages. This evolutionary perspective provides important insights for understanding the flexibility and constraints in developmental programs.
Recent controversies in EMT research often stem from apparent differences in EMT execution across species and contexts. However, an evolutionary perspective reveals that these differences frequently represent variations on conserved themes rather than fundamentally distinct processes. The recognition that EMT encompasses a spectrum of states rather than a simple binary switch has been reinforced by studies across multiple species [9]. This plasticity in EMT execution likely contributed to its evolutionary adaptability, enabling the diversification of body plans while maintaining core morphogenetic principles.
Future research directions should include comparative single-cell analyses of EMT across diverse species to identify deeply conserved gene regulatory networks. The development of new tools for manipulating and visualizing EMT in non-traditional model organisms, particularly those representing key evolutionary transitions, will further illuminate how this fundamental process has shaped animal evolution. Additionally, exploring the intersection between EMT and the evolution of novel structures, such as the neural crest in vertebrates, will provide insights into how conserved processes are redeployed for evolutionary innovation [2] [15].
EMT represents a deeply conserved developmental mechanism that has been central to metazoan evolution from diploblasts to amniotes. The primitive streak of amniote embryos exemplifies how core EMT programs can be adaptively deployed in novel morphological contexts. Conservation of signaling pathways, transcription factors, and cellular processes across vast evolutionary distances highlights the fundamental nature of EMT in animal development. Understanding these evolutionarily conserved principles provides not only insights into developmental biology but also important context for investigating EMT-related pathologies in human disease, particularly cancer and fibrosis. The continued integration of evolutionary perspectives with mechanistic studies will undoubtedly yield new insights into this fundamental biological process.
The formation of the primitive streak is a pivotal event in amniote embryogenesis, establishing the bilateral body plan and initiating the generation of the three germ layers through the process of epithelial-to-mesenchymal transition (EMT). This whitepaper delineates the core signaling pathways—Wnt, Nodal/Vg1, and FGF—that collectively govern this process. We provide a technical analysis of their individual functions, synergistic interactions, and the downstream transcriptional networks that lead to the ingression of epiblast cells. The content is structured to serve researchers and drug development professionals by integrating current molecular insights with practical experimental methodologies, including detailed protocols, essential research reagents, and quantitative data summaries, all framed within the context of EMT-driven primitive streak formation.
The primitive streak is a transient embryonic structure that forms on the epiblast epithelium, serving as the conduit through which cells undergo EMT to internalize and form the mesoderm and endoderm [2]. The molecular core of this transformation is a signaling triad composed of the Wnt, Nodal/Vg1 (TGF-β), and FGF pathways. These pathways are not merely active simultaneously; they function in a highly coordinated, hierarchical network to render epiblast cells competent to respond to inductive signals, dissolve their epithelial adhesions, and acquire a migratory mesenchymal phenotype [2]. Dysregulation of these pathways is implicated in congenital malformations and the pathological reactivation of EMT in cancer and fibrosis, underscoring the importance of their precise understanding for therapeutic development [2] [16].
Core Signaling Pathways and Their Mechanisms
The induction of EMT at the primitive streak is directed by a cascade of signaling events. The table below summarizes the key components and primary functions of each pathway.
Table 1: Core Signaling Pathways in Primitive Streak Formation and EMT
| Pathway | Key Ligands & Receptors | Primary Downstream Effectors | Core Functions in Primitive Streak/EMT |
|---|---|---|---|
| Wnt/β-catenin | Wnt3, β-catenin | TCF/LEF transcription factors | Establishes posterior identity; induces formation of the primitive streak; primes cells for EMT; collaborates with SMAD2/3 [2] [17]. |
| Nodal/Vg1 (TGF-β) | Nodal, Vg1 (Gdf1/Gdf3), Activin, TGF-β | SMAD2/SMAD3 | Specifies mesendoderm; induces EMT-ingression; forms functional heterodimers with Vg1 for potent signaling [2] [18] [19]. |
| FGF | FGF4, FGFR1 | Ras/MAPK, PI3K/AKT | Sustains the EMT regulatory network; promotes cell proliferation and survival post-ingression; maintains mesodermal identity [2] [20]. |
The Wnt/β-catenin Pathway
The Wnt/β-catenin pathway acts as the initial trigger. In the posterior epiblast, canonical Wnt signaling (e.g., via Wnt3) stabilizes β-catenin, allowing its nuclear translocation [2] [17]. Nuclear β-catenin then binds to regulatory regions of primitive streak-specific genes, activating their transcription. A pivotal mechanism is its collaborative interaction with the Nodal effectors SMAD2 and SMAD3. β-catenin occupies DNA alongside SMAD2/3, and this direct interaction is required for the full activation of mesendodermal genes [17]. Furthermore, β-catenin interacts with the pluripotency factor OCT4 at primitive streak enhancers, ensuring that the EMT program is initiated in the appropriate embryonic context [17].
The Nodal/Vg1 (TGF-β) Pathway
Nodal is the key TGF-β ligand responsible for mesendoderm induction. A critical advancement is the understanding that its activity is potentiated by Vg1 (Gdf1/Gdf3). Maternal Vg1 is ubiquitous but inactive when alone, residing in the endoplasmic reticulum. Zygotic Nodal, though localized, is insufficient for robust signaling. They form functional Nodal-Vg1 heterodimers, which are efficiently processed, secreted, and represent the primary bioactive ligands for mesendoderm induction [18]. This pathway signals through receptors that recruit and phosphorylate SMAD2/SMAD3, which then translocate to the nucleus to activate transcription of EMT-related genes, often in concert with β-catenin [17] [19].
The FGF Signaling Pathway
Following the initiation by Wnt and Nodal/Vg1, the FGF pathway is critical for the execution and maintenance of EMT. FGF4 is a major ligand expressed in the primitive streak and signals through receptors like FGFR1 [2] [21]. Its primary role is to maintain the mesenchymal state of the ingressed cells. FGF signaling activates the Ras/MAPK and PI3K/AKT cascades, which promote cell proliferation, survival, and migration [20] [16]. In FGFR1-deficient mice, the primitive streak forms and cells begin to ingress, but their production is not sustained, highlighting FGF's role in maintaining the EMT network rather than initiating it [2].
The following diagram illustrates the sequential interactions and crosstalk between these core pathways in the context of initiating EMT at the primitive streak.
Experimental Analysis of Pathway Interactions
Studying the hierarchical and synergistic relationships between these pathways requires precise experimental models. The following workflow details a standard protocol for manipulating these pathways in a chick embryo model, a classic system for studying primitive streak formation [19].
Table 2: Key Research Reagents for Pathway Manipulation
| Reagent / Tool | Type | Function in Experiment | Example Use Case |
|---|---|---|---|
| CHIR99021 | Small molecule inhibitor | Activates Wnt signaling by inhibiting GSK-3β, stabilizing β-catenin [20] [22]. | Used to test sufficiency of Wnt pathway to induce primitive streak genes. |
| XAV939 | Small molecule inhibitor | Inhibits Wnt signaling by stabilizing Axin, promoting β-catenin degradation [23]. | Used to test necessity of Wnt signaling in streak formation. |
| Cripto (CFC) | Expression plasmid | Overexpression acts as a Nodal/Vg1 pathway co-receptor, potentiating signaling [19]. | Used to test sufficiency of Nodal-like signaling in IM specification. |
| Cerberus-short (Cer-S) | Secreted protein | Specific antagonist of Nodal (but not Activin) signaling [19]. | Used to inhibit Nodal-like signaling specifically to assess its requirement. |
| Dorsalin-Vg1 | Expression plasmid | Constitutively active form of Vg1 used to activate the pathway [19]. | Used to mimic sustained Vg1 signaling and assess its effects on mesoderm. |
| FGF4-/-, FGFR1-/- | Genetic model | Mouse knockout models to study loss of function [2] [21]. | Reveals FGF's role in maintaining mesoderm production post-ingression. |
| Anti-pSmad1/5/8 | Antibody | Detects active BMP signaling via immunohistochemistry [19]. | Used to investigate crosstalk between Nodal and BMP pathways. |
Detailed Protocol: Manipulating Pathways in Chick Embryo
This protocol is adapted from methods used to investigate the role of Vg1/Nodal signaling in intermediate mesoderm specification [19].
Step 1: Embryo Preparation
- Fertilized chick eggs are incubated at 38°C to the desired stage (e.g., HH stage 3-5, corresponding to early gastrulation). The embryo is exposed by creating a window in the eggshell and using thin filters for stability.
Step 2: Experimental Manipulation
- DNA/Reagent Delivery: Plasmid DNA (e.g., pMES-Cripto for gain-of-function, pMES-Cerberus-short for loss-of-function) is mixed with a transfection reagent (e.g., SuperFect) and a fast-green tracking dye. The solution is applied to the embryo. In ovo electroporation is performed using platinum electrodes and a square-wave electroporator (e.g., 5 pulses of 10V, 50ms duration, 100ms interval) to deliver the constructs into the posterior epiblast/capital0 primitive streak region.
- Small Molecule Application: Alternatively, small molecule agonists/antagonists can be applied directly. For instance, the Wnt activator CHIR99021 (10 μM) or inhibitor DKK1 (1 μg/ml) can be soaked into agarose beads and implanted in the embryo's vicinity [22].
Step 3: Post-Manipulation Incubation & Analysis
- The window in the egg is sealed with tape, and the embryo is returned to the incubator for 18-24 hours to develop to the desired stage for analysis.
- Downstream Analysis:
- Whole-mount in situ hybridization (WMISH): Embryos are fixed and hybridized with digoxigenin-labeled RNA probes for primitive streak or mesodermal markers (e.g., Osr1, Pax2 for intermediate mesoderm; Tbx6L for paraxial mesoderm) [19].
- Immunofluorescence (IF): Embryos are cryosectioned and stained with primary antibodies (e.g., anti-Pax2, anti-Lim1, anti-Pax7) and fluorescent secondary antibodies. Nuclear counterstaining with DAPI is performed to visualize tissue architecture [19].
- Phospho-Smad Analysis: To investigate BMP pathway crosstalk, immunofluorescence with an anti-pSmad1/5/8 antibody is performed on whole-mount embryos or sections [19].
Quantitative Data Synthesis
The functional outcomes of pathway manipulation can be quantified by analyzing changes in gene expression domains. The following table synthesizes typical experimental data from gain-of-function (GOF) and loss-of-function (LOF) studies in model organisms.
Table 3: Phenotypic Outcomes of Pathway Manipulation on Mesoderm Specification
| Pathway Manipulation | Experimental Model | Effect on Primitive Streak | Effect on Mesoderm Markers | Key Molecular Readouts |
|---|---|---|---|---|
| Wnt GOF | Mouse embryo | Ectopic primitive streak formation [2]. | Expansion of mesodermal markers. | ↑ Brachyury, ↑ Nodal [2]. |
| Wnt LOF (Wnt3-/-) | Mouse embryo | Absence of primitive streak; failed gastrulation [2]. | Absence of mesoderm. | Loss of Brachyury; no mesoderm formation [2]. |
| Nodal/Vg1 GOF | Chick embryo | Expansion of IM domain at expense of PM [19]. | ↑ Osr1, ↑ Pax2 (IM); ↓ Paraxis (PM) [19]. | Requires active BMP signaling (pSmad1/5/8+) [19]. |
| Nodal/Vg1 LOF (vg1-/-) | Zebrafish embryo | Loss of endoderm and trunk mesoderm [18]. | Loss of ntl (mesoderm), sox32 (endoderm). | Phenocopies Nodal mutants; inactive Nodal-Vg1 heterodimers [18]. |
| FGF LOF (FGFR1-/-) | Mouse embryo | Primitive streak forms but mesoderm production is arrested [2]. | Initial mesoderm markers present but not maintained. | ↓ Mesodermal proliferation and survival [2] [21]. |
| FGF LOF (FGF4-/-) | Mouse embryo | Peri-implantation lethality; defective ICM [21]. | n/a (pre-gastrulation lethality). | Failure in inner cell mass proliferation [21]. |
Integrated Signaling Network and Crosstalk
The pathways do not operate in isolation but are nodes in a tightly regulated network. The following diagram synthesizes the core signaling logic and critical crosstalk mechanisms that ensure robust primitive streak formation.
The integrated signaling network operates on several key principles of crosstalk:
- Hierarchical Initiation: Wnt signaling is a master upstream regulator that establishes the territory competent to form the streak and induces the expression of Nodal [2].
- Ligand Cooperation: Nodal and Vg1 form obligate heterodimers, representing the primary bioactive ligand for mesendoderm induction, explaining why the loss of one disrupts the entire pathway [18].
- Transcription Factor Collaboration: The effector proteins β-catenin and SMAD2/3 do not work in parallel but form a direct complex on the genomic regulatory elements of primitive streak genes, creating an enhanceosome that drives robust transcription [17].
- Synergy with Other Pathways: Nodal-like signaling can induce phosphorylation of BMP pathway effectors (SMAD1/5/8), and active BMP signaling is required for Nodal-mediated IM induction, demonstrating deep pathway integration [19].
- Maintenance and Execution: The FGF pathway, potentially activated downstream of the initial Wnt/Nodal cascade, sustains the mesenchymal state by activating MAPK and other kinase cascades, preventing premature MET and ensuring cell migration [2] [20].
The choreography of Wnt, Nodal/Vg1, and FGF signaling is fundamental to launching the EMT program that creates the primitive streak and establishes the mammalian body plan. The data and protocols presented herein provide a framework for ongoing research into the nuances of this process. A deep understanding of these pathways, their interactions, and their context-dependent effects is not only crucial for developmental biology but also for informing therapeutic strategies against diseases like cancer and fibrosis, where the embryonic EMT program is pathologically reactivated. Future work will continue to elucidate the precise epigenetic and post-translational mechanisms that refine this core signaling network.
Epithelial-to-mesenchymal transition (EMT) is a fundamental biological process critical for embryonic development, including primitive streak formation during gastrulation, and cancer metastasis. This whitepaper provides an in-depth technical analysis of two core EMT-transcribing factors (EMT-TFs), SNAI and TWIST, focusing on their molecular structures, regulatory mechanisms, downstream targets, and experimental approaches for their investigation. Within the context of primitive streak research, we delineate the distinct roles these factors play in germ layer segregation, particularly highlighting how SNAI1 drives classical EMT for mesoderm formation while endoderm specification occurs through SNAI1-independent mechanisms. This resource is designed to equip researchers and drug development professionals with the comprehensive molecular toolkit necessary to advance studies in developmental biology and cancer therapeutics.
The primitive streak is a transient embryonic structure that emerges during gastrulation and serves as the conduit through which epiblast cells ingress to form the mesoderm and definitive endoderm. This massive cellular reorganization is orchestrated by EMT, a process wherein stationary epithelial cells dissolve cell-cell junctions, remodel their cytoskeleton, and acquire motile mesenchymal characteristics. In the mammalian embryo, the formation of the primitive streak establishes the body axes and generates the three germ layers. Type I EMT, occurring during embryogenesis, is distinct from the Type II EMT associated with wound healing and fibrosis, and Type III EMT linked to cancer metastasis [24] [25]. A paradigm-shifting study using mouse embryonic stem cells and mouse embryo knock-in reporter systems demonstrated that germ layer formation involves distinct morphogenetic programs: while the mesoderm is formed by classical, SNAI1-dependent EMT, the definitive endoderm is formed independently of SNAI1 via mechanisms of epithelial cell plasticity [26]. This divergence underscores the sophisticated regulatory landscape governing cell fate during this critical developmental window.
The SNAI Family of Transcription Factors
Molecular Structure and Function
SNAI1 (Snail) is a zinc-finger transcriptional repressor. Its protein structure comprises an N-terminal SNAG domain, a serine-rich domain, a nuclear export signal, and four C-terminal C2H2 zinc-finger domains [24] [25]. Repression of target genes is achieved through the zinc-finger domains binding to E-box sequences (CANNTG) in gene promoter regions. A primary target is CDH1, the gene encoding E-cadherin, a master regulator of epithelial adhesion [24]. The SNAG domain is critical for recruiting chromatin-modifying complexes to enforce transcriptional repression.
Role in the Primitive Streak and Downstream Signaling
In the primitive streak, SNAI1 is a key mediator of EMT for ingressing mesodermal cells. In vertebrates, fibroblast growth factor (FGF)-induced SNAI1 expression contributes to mesoderm cell fate and somite formation [24] [25]. SNAI1 represses E-cadherin, which not only loosens cell adhesion but also increases the cytoplasmic pool of β-catenin, thereby priming the Wnt signaling pathway and reinforcing the EMT program [25].
Beyond development, SNAI1-driven EMT in cancer models confers chemoresistance and cellular plasticity. In thymic epithelial tumors, SNAI1 was identified as a central oncogenic hub that promotes EMT, stemness, and invasion through the PIK3R2/p-EPHA2/GSK3β/β-catenin signaling axis [27]. Similarly, in mammary epithelial cells, SNAI1 overexpression leads to a dramatic reconfiguration of cell death and stem cell maintenance pathways, resulting in a CD44+/CD24- stem-like phenotype and enhanced mammosphere formation [28]. The downstream pathways identified in cancer models often recapitulate developmental signaling, providing insights into the potent capabilities of this transcription factor.
Table 1: Key Downstream Pathways and Functional Outcomes of SNAI1
| Downstream Pathway/Target | Molecular Effect | Functional Outcome | Context |
|---|---|---|---|
| CDH1 (E-cadherin) | Repression | Loss of cell adhesion, increased motility | Development [25], Cancer [24] [27] |
| PIK3R2/p-EPHA2 | Activation of β-catenin | Enhanced stemness and tumorigenesis | Thymic Cancer [27] |
| WNT/β-catenin | Pathway activation | Stem cell maintenance | Cancer Stem Cells [28] |
| NF-κB/MAPK | Pathway activation | Chemoresistance, IL6/IL8 induction | Mammary Epithelial Cells [28] |
The TWIST Family of Transcription Factors
Molecular Structure and Regulatory Mechanisms
TWIST1 is a basic helix-loop-helix (bHLH) transcription factor essential for mesoderm and craniofacial development. Its function is critically regulated by post-translational modifications, particularly acetylation. The diacetylation status of TWIST1 at lysine residues K73 and K76 dictates its interaction with specific chromatin-modifying complexes [29]:
- Non-acetylated TWIST1-K73/76: Selectively interacts with and recruits the NuRD complex (containing HDAC1/2), leading to repression of epithelial target genes like CDH1 and ESR1.
- Diacetylated TWIST1-acK73/76: Binds BRD8, a component of the NuA4/TIP60 histone acetyltransferase complex, leading to activation of mesenchymal genes and MYC.
This dual functionality allows TWIST1 to act as a molecular switch, both repressing the epithelial program and activating the mesenchymal program.
Role in Development and Cancer
During development, TWIST1 is vital for mesoderm formation and its homozygous knockout in mice results in embryonic lethality due to cranial neural tube defects [29] [26]. In cancer, TWIST1 is a major driver of metastasis, stemness, and therapy resistance. For instance, in oncogene-driven lung cancer, TWIST1 was identified as a critical downstream target of the HGF/MET pathway, where it stabilizes the TWIST1 protein and drives acquired resistance to tyrosine kinase inhibitors by suppressing p27 expression [30]. Furthermore, in colon cancer, TWIST1-induced EMT and the acquisition of stem cell-like properties occur via AKT signaling pathways. The specific pathways activated, however, can depend on the cellular context, such as the microsatellite instability (MSI) status of the cells [31].
Table 2: Key Downstream Pathways and Functional Outcomes of TWIST1
| Downstream Pathway/Target | Molecular Effect | Functional Outcome | Context |
|---|---|---|---|
| NuRD Complex | Repression of CDH1, ESR1 | Loss of epithelial features, metastasis | Breast Cancer [29] |
| TIP60 Complex (via BRD8) | Activation of MYC, mesenchymal genes | Enhanced proliferation, metastasis | Breast Cancer [29] |
| AKT/GSK-3β/β-catenin | Nuclear translocation of β-catenin | Stemness (CD44, CD166 expression) | Colon Cancer [31] |
| AKT/NF-κB | Nuclear translocation of NF-κB | Stemness, enhanced invasiveness | Colon Cancer (MSS) [31] |
| p27 | Suppression of p27 | TKI Resistance | Lung Cancer [30] |
Comparative Analysis of SNAI and TWIST in Primitive Streak Formation
The role of EMT-TFs in the primitive streak is highly coordinated and cell-type-specific. High-resolution single-cell RNA sequencing of mouse embryos has revealed a clear lineage bifurcation during gastrulation [26]. Cells committing to the mesoderm lineage strongly upregulate a full EMT program, including high expression of Snail1, Twist1, and other EMT-TFs, and undergo a classic E-cadherin to N-cadherin switch. In stark contrast, cells committing to the definitive endoderm lineage downregulate EMT-TFs like Snail1 and maintain E-cadherin expression while also upregulating N-cadherin, a state known as partial EMT or epithelial cell plasticity [26]. This process is guarded by the transcription factor Foxa2, which acts as an EMT suppressor, shielding the endoderm from a complete mesenchymal transition. Therefore, within the same embryonic structure, SNAI1 is indispensable for mesoderm formation, while endoderm formation proceeds via a distinct, SNAI1-independent pathway.
Experimental Protocols for EMT-TF Research
Identifying EMT-TF Targets: CUT&Tag and RNA-seq
A comprehensive study on thymic epithelial tumors provides a robust methodology for identifying direct downstream targets of SNAI1 [27].
Workflow:
- Cell Line Engineering: Establish stable SNAI1-overexpressing and control cell lines using lentiviral transduction with pSLenti-EF1-CMV-EGFP-P2A-Puro-WPRE vectors.
- Target Identification: Perform CUT&Tag (Cleavage Under Targets and Tagmentation) assay using an anti-SNAI1 antibody to genome-widely profile its DNA binding sites.
- Transcriptional Profiling: Conduct RNA-sequencing on the same models to identify genes with altered expression upon SNAI1 overexpression.
- Data Integration: Intersect CUT&Tag and RNA-seq data to pinpoint direct transcriptional targets of SNAI1 (e.g., PIK3R2).
- Functional Validation:
- Chromatin Immunoprecipitation (ChIP) & CUT&RUN-qPCR: Validate SNAI1 binding to the promoter of the target gene.
- Luciferase Reporter Assay: Confirm that SNAI1 binding directly regulates promoter activity.
- Co-immunoprecipitation (Co-IP) and Phosphoproteomics: Elucidate the functional protein interactions and signaling pathway activation downstream of the identified target.
Lineage Tracing and Single-Cell Analysis in Development
To resolve the morphogenetic programs of germ layer segregation, a combination of lineage labeling and high-resolution transcriptomics is required [26].
Workflow:
- Reporter System Generation: Create knock-in reporter mouse embryonic stem cells or embryos (e.g., Foxa2-Venus, Sox17-mCherry, T-GFP) for specific lineage labeling.
- Time-Resolved Differentiation & Sampling: Differentiate mESCs in a stepwise manner towards mesoderm and endoderm or collect embryos at precise developmental stages (e.g., early-, mid-, late-streak).
- Cell Sorting: Use fluorescence-activated cell sorting (FACS) to isolate distinct progenitor and differentiated cell populations based on reporter expression and surface markers (e.g., CD24).
- Single-Cell RNA Sequencing (scRNA-seq): Perform high-throughput scRNA-seq on the sorted populations.
- Computational Analysis:
- Pseudotime and Trajectory Inference: Use algorithms like scVelo and CellRank to map the sequence of transcriptional changes and predict cell fate.
- Lineage Driver Genes: Identify key genes that drive the separation between mesoderm and endoderm lineages.
- EMT Program Scoring: Analyze the expression dynamics of EMT-TFs (SNAI1, TWIST1, ZEB1/2) and adhesion molecules (CDH1, CDH2) across the trajectories.
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Models for Investigating SNAI and TWIST
| Reagent / Model | Specific Example | Function in Research |
|---|---|---|
| Reporter Cell Lines | Foxa2-Venus; Sox17-mCherry; T-GFP mESCs [26] | Lineage tracing and live-cell tracking during germ layer specification. |
| Plasmids for Overexpression | pSLenti-EF1-CMV-EGFP-P2A-Puro-WPRE [27] | Stable, selectable overexpression of genes of interest (e.g., SNAI1, TWIST1). |
| Plasmids for Knockdown | pSLenti-U6-CMV-EGFP-F2A-Puro-WPRE shRNA [27] | Stable, selectable knockdown of target genes via short hairpin RNA. |
| Small Molecule Inhibitors | MET Tyrosine Kinase Inhibitors [30], Quinacrine (NF-κB) [31] | Functional validation of signaling pathways and therapeutic potential. |
| Pathway Reporter Assays | TCF/β-catenin Luciferase, 17x4-TK-Luc [29] [28] | Quantifying activity of specific signaling pathways downstream of EMT-TFs. |
| Antibodies for Functional Assays | Anti-SNAI1 (for CUT&Tag) [27], Anti-active Caspase-3 (apoptosis) [28], Anti-CD44/CD24 (FACS) [28] | Detecting protein localization, expression, and cell surface phenotypes. |
SNAI and TWIST are master regulators of EMT with distinct yet complementary roles in development and disease. The primitive streak serves as a powerful model to deconstruct their functions: SNAI1 is a non-redundant driver of classical EMT for mesoderm formation, whereas TWIST1's context-dependent transcriptional output fine-tunes cell fate. The experimental frameworks and tools outlined herein provide a roadmap for discovering novel downstream effectors and therapeutic targets. Future research exploiting single-cell multi-omics and sophisticated lineage-tracing models will continue to unravel the complex regulatory networks governed by these transcription factors, with profound implications for regenerative medicine and oncology.
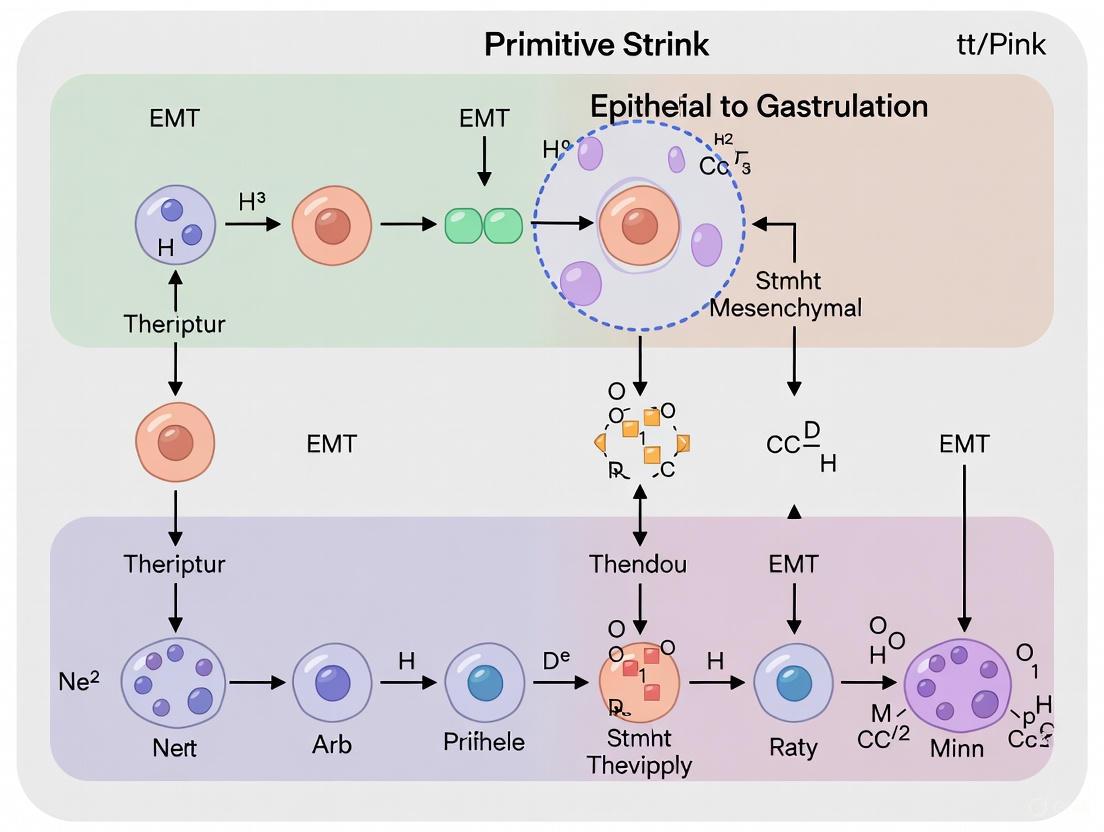
Epithelial-mesenchymal transition (EMT) is a fundamental cellular process during embryonic development, tissue repair, and disease progression. In the context of primitive streak formation in amniote embryos, EMT enables the transformation of polarized epithelial cells from the epiblast into migratory mesenchymal cells that give rise to the mesoderm and definitive endoderm, establishing the three primordial germ layers of the embryo [10] [2]. This transition involves dramatic cellular reorganization, including the loss of apicobasal polarity, dissolution of cell-cell adhesion structures, and acquisition of front-rear polarity with migratory capacity [32] [2]. The primitive streak, a transient structure that forms along the posterior midline of the embryonic disc, serves as the site where EMT is initiated and regulated through complex signaling networks and transcription factor cascades [1] [33]. Understanding the precise cellular and molecular mechanisms governing this transition provides critical insights into normal development and pathological processes, including cancer metastasis and organ fibrosis, where EMT is reactivated [4] [34].
Core Cellular Changes During EMT at the Primitive Streak
The transition from an epithelial to mesenchymal state involves a coordinated sequence of morphological changes that can be delineated into distinct stages. The process begins with cells poised to undergo EMT, followed by the disruption of intercellular adhesion, loss of apical-basal polarity, cytoskeletal reorganization, and ultimately, cell ingression and migration [2].
Table 1: Sequential Cellular Changes During EMT in Primitive Streak Formation
| Stage of EMT | Key Cellular Events | Major Molecular Markers |
|---|---|---|
| Polarized Epithelial Cell | Apicobasal polarity; intact basement membrane; mature cell junctions [2] | E-cadherin, Occludin, ZO-1, Cytokeratins [32] |
| Early EMT Initiation | Loss of basal lamina; apical constriction; reduced cell-cell adhesion [10] [2] | Reduced E-cadherin; Increased Snail1/2 [32] [2] |
| Intermediate/Hybrid State | Partial epithelial/mesenchymal features; may retain some contacts [35] [4] | Co-expression of epithelial and mesenchymal markers [34] |
| Full Mesenchymal Phenotype | Spindle-shaped morphology; front-rear polarity; migratory capacity [2] [34] | Vimentin, N-cadherin, Fibronectin, α-SMA [32] |
In the mouse embryo, studies using live 4-dimensional imaging and immunohistochemistry have revealed that the primitive streak develops in situ through progressive initiation of EMT in the posterior epiblast, rather than through large-scale migration or convergence of a cell population [10]. The loss of the basal lamina represents the initial step of this EMT process and is tightly correlated with the ingression of nascent mesoderm. Once the basal lamina is disrupted in a specific region, cells exit the epiblast through apical constriction to enter the primitive streak [10]. This mechanism appears distinct from other amniote models like chick and rabbit, where extensive cell rearrangements contribute to streak formation [33].
Signaling Pathways Regulating EMT in the Primitive Streak
The initiation and progression of EMT at the primitive streak are governed by a complex interplay of multiple conserved signaling pathways. These pathways integrate spatial and temporal cues from the embryonic microenvironment to activate transcription factors that execute the EMT program.
Diagram 1: Signaling pathways and transcriptional regulation of EMT at the primitive streak. The diagram illustrates how inductive signals (BMP, Wnt, Nodal, FGF) are integrated to activate a hierarchy of EMT transcription factors, ultimately leading to cellular changes that enable ingression. The anterior visceral endoderm (AVE) secretes inhibitors that restrict these signals to the posterior region.
Key Signaling Interactions
Wnt/β-catenin Signaling: Wnt signaling, particularly through Wnt3, is a primary initiator that renders epiblast cells competent to undergo EMT [2] [33]. Mouse embryos deficient in Wnt3 fail to form a primitive streak, while overexpression of Wnt8c leads to multiple primitive streaks, demonstrating its crucial role [2]. Wnt signaling activates the stabilization and nuclear translocation of β-catenin, which initiates the expression of primitive streak genes [33].
TGF-β Superfamily (Nodal and Vg1): Nodal and Vg1 (a TGF-β family member) cooperate with Wnt to induce primitive streak formation [2]. Ectopic expression of Vg1 alone can induce an additional primitive streak in chick embryos [2]. Nodal mutants fail to complete gastrulation, highlighting its necessity in this process [2]. The anterior visceral endoderm (AVE) and hypoblast secrete inhibitors of Nodal (such as Cerberus and Lefty1), which restrict streak formation to the posterior end [1] [33].
BMP and FGF Signaling: BMP4 secreted from the extraembryonic ectoderm (ExE) works in a gradient with higher signaling in the posterior, promoting mesoderm formation [33]. FGF signaling is thought to work with Nodal to maintain the EMT regulatory network, as FGFR1-deficient mice show arrested production of mesodermal cells after initial streak formation [2].
Transcriptional Control of EMT
The signaling pathways described above converge on a core set of EMT-inducing transcription factors (EMT-TFs) that execute the cellular reprogramming.
Hierarchy of EMT Transcription Factors
Snail Family (Snail1 and Snail2/Slug): Snail1 acts as a pioneer factor that is rapidly activated in response to EMT-inducing signals [34]. It initiates the EMT program by repressing the transcription of E-cadherin (CDH1) and other epithelial genes [32] [2]. Studies in mammalian cell models show that SNAIL1 exhibits two-wave dynamics, with an initial fast burst followed by recruitment of other EMT-TFs [34].
Twist, ZEB, and PRRX Families: Following the initial Snail activation, other transcription factors including Twist, ZEB1/2, and PRRX1 are recruited to reinforce the mesenchymal phenotype [32] [34]. PRRX1 is particularly important for driving the invasive mesenchymal trajectory and is required for the full completion of EMT in some contexts [34]. Knockdown of PRRX1 prevents full EMT and reverts cells to a partial EMT state [34].
Table 2: Key EMT Transcription Factors and Their Functions
| Transcription Factor | Primary Function in EMT | Regulation in Primitive Streak |
|---|---|---|
| Snail1/Snail2 | Pioneer factors; repress E-cadherin transcription; initiate EMT program [32] [34] | Activated by Wnt, Nodal, and FGF signaling; first wave of EMT-TF expression [2] [34] |
| Twist1/Twist2 | bHLH factors; promote mesenchymal gene expression; cooperate with Snail [32] | Second wave of expression; promotes progression to full EMT [34] |
| ZEB1/ZEB2 | Transcriptional repressors of epithelial genes; promote mesenchymal phenotype [32] | Activated downstream of initial EMT signals; reinforce EMT program [32] |
| PRRX1 | Drives invasive mesenchymal trajectory; required for full EMT in some contexts [34] | Late-stage EMT-TF; associated with migratory capacity and metastatic potential [34] |
Species-Specific Variations in Primitive Streak Formation
While the core program of EMT is conserved across amniotes, significant differences exist in the cellular behaviors that form the primitive streak in different model organisms.
Table 3: Comparative Mechanisms of Primitive Streak Formation
| Species | Embryo Morphology | Cellular Mechanisms of Primitive Streak Formation | Key References |
|---|---|---|---|
| Mouse | Cup-shaped | Primitive streak forms in situ by progressive EMT without large-scale cell migration or convergence-extension [10] [33] | [10] |
| Chick | Flat disc | "Polonaise movements" - large-scale coordinated cell flows with convergence-extension at the midline [33] [36] | [33] [36] |
| Rabbit | Flat disc | L- and U-shaped cell movement patterns from peripheral regions toward the primitive streak [33] | [33] |
These differences may stem from the vast differences in size and morphology of the embryos at the time of gastrulation. Chick and rabbit embryos are many times larger than the mouse embryo and exist as flat discs, while the mouse embryo is cup-shaped [33].
Experimental Approaches for Studying EMT in Primitive Streak
Key Methodologies
Advanced imaging and molecular techniques have been crucial for elucidating the mechanisms of EMT in primitive streak formation.
Live 4-Dimensional Imaging: This approach allows for the dynamic observation of cell behaviors during primitive streak formation in real-time. In mouse embryos, this technique revealed the absence of large-scale cell migration, supporting the in situ formation model [10].
Immunohistochemistry and Electron Microscopy: These methods enable the visualization of cellular structures during EMT, including the loss of basal lamina, disruption of cell junctions, and cytoskeletal reorganization [10] [4].
Single-Cell RNA Sequencing: Recent applications of scRNA-seq in chick embryos have revealed the cellular composition of Hensen's node (the avian organizer), identifying distinct anterior and posterior cell populations with different transcriptional profiles and inductive properties [36].
Genetic Manipulation: The use of knockout mice for specific signaling components (e.g., Wnt3, Nodal, FGFR1) has been instrumental in establishing the functional requirements of these pathways in primitive streak formation and EMT [2] [33].
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Reagents for Studying EMT in Primitive Streak Research
| Reagent/Category | Specific Examples | Function/Application | Key References |
|---|---|---|---|
| Signaling Agonists | TGF-β1, BMP4, FGF2, Wnt3a, Activin A | Induce EMT and primitive streak formation in model systems [33] [34] | [33] [34] |
| Signaling Inhibitors | SU5402 (FGF inhibitor), SB-431542 (Nodal/Activin inhibitor), NOG (BMP inhibitor) | Block specific signaling pathways to assess function in EMT [33] [34] | [33] [34] |
| EMT Marker Antibodies | E-cadherin, N-cadherin, Vimentin, ZO-1, Snail1, Fibronectin | Detect epithelial and mesenchymal protein expression by IHC/IF [10] [32] | [10] [32] |
| Genetic Tools | siRNA/shRNA (e.g., anti-SNAIL1, anti-PRRX1), CRISPR-Cas9, Transgenic mice | Knockdown or knockout specific genes to study function in EMT [34] | [34] |
| 3D Culture Matrices | Collagen matrices, Matrigel | Study invasive behavior in a three-dimensional environment [34] | [34] |
Diagram 2: Experimental approaches in primitive streak EMT research. The diagram connects key methodologies (blue nodes) with the specific biological insights (green nodes) they provide in the study of EMT during primitive streak formation.
The process of EMT at the primitive streak represents a paradigm of coordinated cellular reprogramming essential for metazoan development. The transformation from apicobasal polarized epithelial cells to migratory mesenchymal cells involves a precisely orchestrated sequence of cellular events, including the loss of cell adhesion, cytoskeletal reorganization, and acquisition of migratory capacity. This process is regulated by an evolutionarily conserved network of signaling pathways—Wnt, Nodal/TGF-β, BMP, and FGF—that activate a hierarchical cascade of transcription factors, with Snail proteins acting as pioneer factors and PRRX1 driving the invasive mesenchymal trajectory. While the core EMT program is conserved, species-specific variations in primitive streak formation highlight the adaptability of this fundamental process. Ongoing research using advanced technologies like single-cell transcriptomics and high-resolution live imaging continues to refine our understanding of EMT heterogeneity and dynamics, with significant implications for understanding developmental disorders, tissue regeneration, and cancer metastasis.
Modern Techniques and Models for Analyzing Primitive Streak EMT
Epithelial-mesenchymal transition (EMT) is a fundamental evolutionary conserved process wherein epithelial cells lose apical-basal polarity and cell-cell adhesion and acquire migratory mesenchymal properties. Its reverse process, mesenchymal-epithelial transition (MET), similarly underpins critical developmental events [4]. The primitive streak, a transient embryonic structure that forms during gastrulation, serves as the quintessential model for studying EMT in vivo [37] [2]. In amniotes (mammals, birds, and reptiles), the primitive streak is the site where epiblast cells undergo EMT, ingress, and migrate to form the mesoderm and definitive endoderm, thereby establishing the three germ layers [37] [2]. Research into the mechanisms of EMT, using the primitive streak as a paradigm, is therefore crucial not only for understanding embryonic development but also for elucidating the mechanisms underlying cancer metastasis, organ fibrosis, and wound healing in adult tissues [2] [4] [14]. This whitepaper provides a comprehensive technical guide to the major in vivo model systems—murine, avian, and non-traditional embryos—used in primitive streak and EMT research, detailing their unique advantages, key experimental findings, and essential methodologies.
Comparative Analysis of Murine and Avian Model Systems
Mouse and chicken embryos are the predominant models for studying gastrulation and EMT due to their well-characterized development and amenability to genetic manipulation. The table below summarizes their core characteristics and applications.
Table 1: Core Characteristics of Murine and Avian Embryo Models for EMT Research
| Feature | Murine Model (M. musculus) | Avian Model (G. gallus / Q. coturnix) |
|---|---|---|
| Embryonic Development | In utero, requiring precise developmental staging [37]. | Ex utero in easily accessible eggs, facilitating observation. |
| Key EMT Findings | Primitive streak forms in situ via progressive EMT without large-scale cell migration; distinct EMT programs for endoderm (Snail1-independent) and mesoderm [10] [26]. | Classic model for defining EMT; exhibits "partial EMT" or "metastable phenotype" during epiboly [2] [38]. |
| Key Signaling Pathways | Wnt, Nodal (TGF-β superfamily), and FGF signaling are critical for inducing and maintaining EMT [2]. | Wnt, Vg1 (TGF-β superfamily), and FGF drive primitive streak formation and EMT [2]. |
| Live Imaging Modalities | Explanted embryos cultured ex vivo for time-lapse imaging [10] [14]. | Direct access allows for high-resolution, long-duration live imaging of processes like epiboly [38]. |
| Genetic Tractability | Sophisticated gene targeting (knockouts, conditional alleles, Cre-lox) and transgenic reporter lines (e.g., for Foxa2, Snail1, Brachyury) [26]. | Electroporation for gene overexpression/knockdown; transgenic quail lines [38]. |
| Typical Readouts | scRNA-seq for lineage tracing, immunohistochemistry, analysis of mutant phenotypes [26]. | Immunofluorescence for EMT markers, time-lapse analysis of collective cell migration [38]. |
Murine Models: A Platform for Genetic Dissection of EMT
The mouse embryo offers unparalleled genetic tools, enabling precise lineage tracing and functional studies of EMT regulators.
Key Morphogenetic and Molecular Events
Mouse gastrulation begins at embryonic day (E) 6.25 with the appearance of the primitive streak [37]. Epiblast cells destined to ingress undergo a characteristic sequence of events at the streak:
- Loss of Basal Lamina: The breakdown of the underlying basement membrane is the initial step, closely correlated with subsequent cell ingression [10].
- Apical Constriction and Cell Elongation: Ingressing cells adopt a "bottle-shaped" morphology, narrowing their apical surface while maintaining transient contacts with neighbors [37].
- Junctional Remodeling: Adherens junctions, including those containing E-cadherin, are dismantled. In mesoderm progenitors, this is accompanied by a cadherin switch to N-cadherin [37] [26].
- Cytoskeletal Rearrangement: The actin cytoskeleton is reorganized to facilitate apical constriction and the formation of basal protrusions, enabling cells to delaminate and migrate [37] [14].
Detailed Protocol: Single-Cell RNA Sequencing of EMT Lineages
The integration of high-resolution scRNA-seq with genetic lineage tracing in mice has revolutionized the understanding of germ layer segregation [26].
Workflow:
- Embryo Dissociation: Isolate E6.5-E7.5 mouse embryos expressing a lineage reporter (e.g., Foxa2-Venus) and dissociate the posterior region containing the primitive streak into a single-cell suspension.
- Fluorescence-Activated Cell Sorting (FACS): Sort cells into distinct populations based on reporter fluorescence intensity (e.g., FVFlow epiblast progenitors, FVFhigh transitory progenitors) and surface markers (e.g., CD24) [26].
- Library Preparation and Sequencing: Use a microfluidic platform (e.g., 10x Genomics) to barcode cells and generate sequencing libraries for high-throughput scRNA-seq.
- Bioinformatic Analysis:
- Clustering and Annotation: Perform dimensionality reduction (e.g., UMAP) and cluster cells based on transcriptional similarity. Annotate clusters using known marker genes (e.g., T for mesoderm, Foxa2 for endoderm).
- Trajectory Inference: Utilize algorithms like scVelo and CellRank to compute RNA velocity and fate probabilities, reconstructing the developmental trajectory from epiblast to mesoderm or endoderm [26].
- Differential Expression: Identify genes that are dynamically regulated along each trajectory, revealing key drivers of EMT and MET.
Key Finding: This approach demonstrated that while mesoderm formation involves a classical Snail1-dependent EMT with a clear E- to N-cadherin switch, definitive endoderm formation occurs via a Snail1-independent mechanism of epithelial plasticity, maintaining E-cadherin expression and bypassing a full mesenchymal intermediate [26].
Avian and Non-Traditional Embryo Models
Avian embryos, along with other non-mammalian models, provide exceptional accessibility for direct observation and manipulation.
The Avian Model: Epiboly and Partial EMT
The quail embryo has been re-established as a powerful model for studying collective epithelial migration and partial EMT during epiboly, the process of blastoderm expansion over the yolk [38].
Key Experimental Findings:
- Co-expression of Markers: Blastoderm edge cells robustly co-express the mesenchymal marker vimentin and the epithelial markers E-cadherin, β-catenin, and cytokeratin, defining a partial EMT or "metastable" phenotype [38].
- Non-Proliferative Front: BrDU incorporation assays show that the edge cell population is non-proliferative, with expansion driven by the continuous recruitment of cells from the inner epithelial zone [38].
- Basement Membrane Remodeling: The extracellular matrix protein laminin, a key component of the basement membrane, is absent from the blastoderm edge, consistent with a migratory, mesenchymal-like state [38].
Zebrafish and Drosophila: Insights from Live Imaging
- Zebrafish: Live imaging of gastrulation and neural crest delamination in transparent zebrafish embryos has provided unparalleled views of cytoskeletal dynamics and collective cell migration during EMT [14].
- Drosophila: Studies in fruit fly embryos have revealed intricate details of adherens junction remodeling, demonstrating that the EMT transcription factor Snail and actomyosin contractility drive junctional disassembly through both transcriptional and post-translational mechanisms [14].
Signaling Pathways and Molecular Regulation of EMT
A complex signaling network governs EMT at the primitive streak. The following diagram synthesizes the core pathway across model systems.
The Scientist's Toolkit: Essential Reagents and Models
This table catalogs key research tools for investigating EMT in vivo.
Table 2: Essential Research Reagent Solutions for In Vivo EMT Studies
| Reagent / Model Name | Type | Key Application / Function | Example Use-Case |
|---|---|---|---|
| Foxa2-Venus; Sox17-mCherry mice/mESCs [26] | Dual knock-in reporter | Labels definitive endoderm progenitors and lineage for live tracking and sorting. | Resolving endoderm morphogenesis independent of an EMT-MET cycle [26]. |
| T-GFP; Foxa2-tagRFP mESCs [26] | Dual knock-in reporter | Enables simultaneous resolution of mesoderm (T) and endoderm (Foxa2) segregation in vitro. | scRNA-seq and pseudotime analysis of lineage bifurcation [26]. |
| Fsp1-Cre; Rosa26-RFP-GFP mice [39] | EMT-driven fluorescent switch | In vivo tracing of EMT initiation; RFP+ epithelial cells switch to GFP+ upon Fsp1 activation. | Intravital imaging of EMT dynamics in breast cancer models [39]. |
| Anti-E-cadherin Antibody [37] [26] | Immunohistochemistry Marker | Stains adherens junctions, hallmark of epithelial state; loss is EMT indicator. | Demonstrating maintained E-cadherin in endoderm progenitors vs. loss in mesoderm [26]. |
| Anti-N-cadherin Antibody [37] [26] | Immunohistochemistry Marker | Stains nascent cell-cell contacts in mesenchymal cells; "cadherin switch". | Identifying mesoderm cells undergoing EMT within the primitive streak [37]. |
| Anti-Vimentin Antibody [37] [38] | Immunohistochemistry Marker | Intermediate filament protein, standard marker for mesenchymal cells. | Identifying mesenchymal cells in mouse streak [37] and partial EMT in quail edge cells [38]. |
| Anti-Laminin Antibody [38] | Immunohistochemistry Marker | Labels the basement membrane; breakdown is an early EMT step. | Visualizing basement membrane dissolution at the primitive streak and blastoderm edge [10] [38]. |
| TGF-β (Transforming Growth Factor-β) [34] | Soluble Factor / Inducer | Potent inducer of EMT in multiple cell and embryo contexts. | Treatment of MDCK cells in vitro to dissect invasive vs. non-invasive EMT trajectories [34]. |
Visualizing EMT: Experimental Workflows from Model to Insight
The diagram below outlines a generalized workflow for conducting and analyzing an in vivo EMT experiment.
The processes of gastrulation and the epithelial-to-mesenchymal transition (EMT) represent pivotal events in mammalian development and disease. Gastrulation establishes the fundamental body plan, characterized by the emergence of the three germ layers through the primitive streak, a process inherently linked to EMT. Similarly, EMT, wherein epithelial cells lose polarity and acquire migratory mesenchymal properties, is co-opted in pathologies such as cancer metastasis. Research in these areas has been historically constrained by the inaccessibility of in vivo embryos and the limitations of two-dimensional culture systems. The advent of sophisticated stem cell-based in vitro models has revolutionized this landscape, providing unprecedented tools to dissect these complex events with high reproducibility and scalability. This technical guide details the core methodologies and applications of these advanced models, framing them within the critical context of primitive streak research and EMT.
Core In Vitro Models for Gastrulation and EMT
3D Gastruloids: Capturing Gastrulation Dynamics
3D gastruloids are aggregates of embryonic stem cells that self-organize to recapitulate key spatiotemporal events of post-implantation development [40]. Their tractable nature and scalability make them an unparalleled platform for high-throughput investigation of normal and pathological embryogenesis.
- Core Principle: Embryonic stem cells are aggregated and cultured in 3D, often with specific biochemical cues, to initiate self-organization.
- Key Features: These models reproducibly form structures that mimic the breaking of embryonic symmetry and the emergence of germ layer progenitors, including those expressing markers of the primitive streak, mesoderm, and endoderm.
- Applications: Gastruloids are extensively used to study patterning, the role of specific signaling pathways in development, and the impact of genetic or chemical perturbations.
Totipotent-like Stem Cell Models: From ZGA to Gastrulation
A recent breakthrough involves using chemically induced totipotent-like cells to generate embryo models that recapitulate a continuous developmental trajectory [41].
- Core Principle: Mouse pluripotent stem cells are treated with a defined chemical cocktail to induce a totipotent-like state, which is then leveraged to generate structured embryo models.
- Key Features: This system sequentially mimics mouse embryogenesis from embryonic day 1.5 to 7.5, encompassing zygotic genome activation (ZGA), lineage diversification, blastocyst formation, and post-implantation development into egg cylinders. Notably, these models undergo gastrulation, indicated by the formation of a primitive streak-like structure [41].
- Applications: This model is ideal for studying the complete continuum of early development, particularly the transition from pre- to post-implantation stages and the initial stages of germ layer formation.
Human Stem Cell-Derived Models: Post-Implantation Lineages
For human development, stem cell models that capture post-implantation events are invaluable. One such model is the human extra-embryoid (hEE), which self-patterns from human pluripotent stem cells [42].
- Core Principle: Human pluripotent stem cells are aggregated in 3D and exposed to a spontaneous differentiation medium, prompting self-organization without establishing placental cell types.
- Key Features: The system efficiently differentiates into diverse cell states of the perigastrulating human embryo, including signatures of the post-implantation epiblast, amniotic ectoderm, primitive streak, and early mesoderm [42]. It captures the co-development of embryonic and extra-embryonic hypoblast-like lineages, which are integral to establishing the body plan.
- Applications: This platform is powerful for studying human-specific developmental events, including the initial patterning of amniotic and primitive streak-like lineages.
3D EMT Models in Bioengineered Matrices
To specifically investigate EMT and its reverse process (MET), 3D models using bioengineered matrices have been developed. These models recapitulate the plasticity and phenotypic heterogeneity of cells in transition.
- Core Principle: Normal epithelial cells are encapsulated in a tunable 3D matrix (e.g., RGD-alginate hydrogel) that mimics the tissue microenvironment. EMT is then induced, for example, with TGFβ1 supplementation [43].
- Key Features: This setup allows for the study of intermediate, metastable EMT states—hybrid cell populations that co-express both epithelial and mesenchymal markers. These states are highly relevant for understanding metastatic dissemination and are difficult to capture in vivo [43] [44].
- Applications: This model is optimal for dissecting the role of the extracellular matrix in EMT, screening for drugs that interfere with transition plasticity, and investigating the functional properties of cells in hybrid states.
Table 1: Summary of Key In Vitro Models for Gastrulation and EMT
| Model Type | Stem Cell Origin | Key Developmental Stage Recapitulated | Readouts & Landmarks |
|---|---|---|---|
| 3D Gastruloids [40] | Mouse/Embryonic Stem Cells | Mammalian gastrulation | Primitive streak-like patterning, germ layer specification |
| Totipotent-like Cell Model [41] | Mouse Pluripotent Stem Cells | E1.5 to E7.5 mouse embryogenesis | ZGA, blastocyst formation, primitive streak-like structure |
| Human Extra-embryoid (hEE) [42] | Human Pluripotent Stem Cells | Carnegie stage 4-7 (human post-implantation) | Primitive streak-like, mesoderm, amniotic ectoderm, hypoblast |
| 3D EMT/MET Model [43] | Mammary Epithelial Cells (e.g., EpH4) | EMT and MET dynamics | E-cadherin loss, N-cadherin/vimentin gain, hybrid state markers |
Experimental Protocols for Key Methodologies
Protocol 1: Generation of a Continuous Mouse Embryo Model from Totipotent-like Cells
This protocol is adapted from the study that induced totipotent-like cells to model development from ZGA to gastrulation [41].
Step 1: Induction of Totipotent-like Cells
- Starting Material: Use mouse extended pluripotent stem (EPS) cells.
- Culture Conditions: Seed cells in AggreWell plates under 3D conditions.
- Induction Cocktail: Treat cells with a chemical cocktail containing CD1530 (retinoic acid agonist, 1 µM), PD0325901 (MEK inhibitor, 1 µM), CHIR-99021 (Wnt agonist, 3 µM), and elvitegravir (10 µM). This is designated as Stage 1 (S1).
- Duration: Culture for 48-72 hours. Validate successful induction via upregulation of totipotency markers (e.g., ZSCAN4, MuERV-L) using immunofluorescence.
Step 2: Stepwise In Vitro Development
- Transition: After aggregate formation in S1, transfer structures to a sequential culture system.
- Stage 2 (S2): Culture in a specific medium formulation that supports pre- to post-implantation transition, promoting the formation of a blastocyst-like structure.
- Stage 3 (S3): Further culture in conditions that support the development of post-implantation egg cylinder-like structures.
- Key Milestones: Monitor for spatial organization, formation of embryonic and extra-embryonic-like compartments, and finally, the emergence of a primitive streak-like structure indicating gastrulation.
Step 3: Analysis
- Single-cell RNA-seq: To transcriptomically characterize the different subpopulations and their developmental trajectories.
- Immunofluorescence: For spatial validation of key markers (e.g., SOX2, SOX17, CDX2, TBXT).
- Lineage Tracing: Perform chimeric experiments to confirm the bi-potential contribution of the totipotent-like cells to both embryonic and extra-embryonic tissues in vivo.
Protocol 2: Establishing a 3D EMT/MET Model in Bioengineered Hydrogels
This protocol outlines the creation of a 3D microenvironment to study EMT and its reversion [43].
Step 1: Preparation of the 3D Matrix
- Matrix Composition: Use soft alginate hydrogels (∼1 wt.% to achieve stiffness G' ≈ 200 Pa) functionalized with cell-adhesive RGD peptides (200 µM).
- Encapsulation: Resuspend normal murine mammary epithelial EpH4 cells at a density of 5 × 10^6 cells/mL in the alginate solution.
- Cross-linking: Cross-link the cell-alginate suspension with calcium to form stable hydrogels. Culture in standard medium for 10-14 days to allow for epithelial morphogenesis and the formation of polarized, lumen-containing acini-like structures.
Step 2: Induction of EMT
- Inducer: Supplement the culture medium with 16 ng/mL of TGFβ1.
- Duration: Treat for 7 days.
- Validation: Confirm EMT by assessing morphological change to a spindle-like shape, downregulation of E-cadherin (protein and CDH1 mRNA), and upregulation of mesenchymal markers like N-cadherin (CDH2) and vimentin.
Step 3: Induction of MET to an Intermediate State
- Reversion: Remove TGFβ1 from the culture medium.
- Outcome: This generates a reversed epithelial (RE) cell population, which is a hybrid state co-expressing both epithelial and mesenchymal markers.
- Functional Assays: Assess the increased proliferative and clonogenic activity of these RE cells compared to fully mesenchymal cells.
Table 2: Key Signaling Molecules and Research Reagent Solutions
| Reagent / Molecule | Function / Target | Application in Model Systems |
|---|---|---|
| TGFβ1 [43] [44] | Potent inducer of EMT; key morphogen in gastrulation | Used in 3D EMT models to initiate transition; implicated in primitive streak signaling. |
| CHIR-99021 [41] | GSK-3 inhibitor; activates Wnt/β-catenin signaling | Critical for maintaining totipotent-like state; essential for self-organization in gastruloids. |
| PD0325901 [41] | MEK inhibitor; suppresses ERK/MAPK signaling | Enhances totipotency induction; used in differentiation protocols. |
| CD1530 [41] | Retinoic acid receptor agonist | Key inducer of totipotency from pluripotent states. |
| RGD-Alginate Hydrogel [43] | Synthetic ECM with tunable biochemical/mechanical properties | Provides a defined 3D microenvironment for epithelial morphogenesis and EMT studies. |
| SOX2 / SOX17 Antibodies [42] | Markers for epiblast and hypoblast/endoderm lineages, respectively | Essential for immunostaining to validate lineage specification in human and mouse models. |
Signaling Pathways and Workflow Visualization
The following diagrams, generated using Graphviz, illustrate the core signaling pathways and experimental workflows central to these models. The color palette adheres to the specified guidelines (#4285F4, #EA4335, #FBBC05, #34A853, #FFFFFF, #F1F3F4, #202124, #5F6368), with explicit text coloring for contrast.
Signaling Network in Primitive Streak and EMT
This diagram outlines the core signaling pathways involved in primitive streak formation and the induction of EMT.
Diagram 1: Signaling in Primitive Streak and EMT. Green nodes represent upstream signaling pathways. Red nodes are EMT-transcription factors. Yellow nodes are effector molecules. Blue nodes are the key biological processes. Inhibitory interactions are shown with a bar-headed arrow.
Experimental Workflow for 3D EMT/MET Model
This diagram details the sequential steps for establishing and analyzing the 3D EMT/MET model in bioengineered hydrogels.
Diagram 2: 3D EMT/MET Model Workflow. The process flows from epithelial cell encapsulation, through morphogenesis, TGFβ1-induced EMT, and subsequent MET to a hybrid state upon stimulus removal. Dashed lines indicate analysis points.
Stem cell-based in vitro models have fundamentally transformed our ability to investigate the black box of early mammalian development and associated disease processes. The systems detailed here—from gastruloids and totipotent-like cell models that capture the breadth of gastrulation to sophisticated 3D microenvironments that dissect EMT plasticity—provide a robust, ethical, and scalable toolkit for the scientific community. When framed within the context of primitive streak research, these models offer a mechanistic lens through which to view the initial stages of cell fate determination and the onset of cellular migration. As these technologies continue to evolve, integrating higher complexity and fidelity, they hold immense promise for advancing our understanding of human development, congenital disorders, and the metastatic cascade in cancer.
Epithelial-to-mesenchymal transition (EMT) represents a fundamental biological process driving cellular plasticity during embryonic development, particularly in primitive streak formation during gastrulation. In this critical developmental window, epiblast cells undergo precisely regulated state transitions to give rise to the mesoderm and definitive endoderm germ layers [26]. The molecular profiling of these transitions through transcriptomic, proteomic, and multi-omic approaches has revolutionized our understanding of cell fate decisions, revealing complex regulatory networks and dynamic protein expression changes that coordinate these developmental events.
Contemporary research has demonstrated that germ layer segregation employs distinct morphogenetic programs: while mesoderm formation occurs through classical EMT characterized by Snail1-mediated E-cadherin to N-cadherin switching, definitive endoderm formation proceeds through Snail1-independent mechanisms of epithelial cell plasticity, maintaining E-cadherin while coordinately upregulating N-cadherin [26]. Resolving these differential programs requires sophisticated molecular profiling approaches that can capture both transcriptional and proteomic changes with high resolution across rare cell populations.
The integration of multi-omics data with advanced computational models now enables researchers to reconstruct developmental trajectories, identify novel intermediate cell states, and uncover regulatory mechanisms governing cell fate decisions during primitive streak formation. This technical guide outlines the core methodologies, analytical frameworks, and applications of molecular profiling approaches within this specific developmental context, providing researchers with practical resources for implementing these techniques in their investigations of EMT in gastrulation and cancer progression.
Core Signaling Pathways in EMT and Primitive Streak Formation
Molecular Regulators of EMT
EMT is orchestrated by a core set of transcription factors and signaling pathways that show remarkable conservation between developmental and pathological contexts. Key transcription factors include Snail, Slug, Twist, and ZEB1/2, which collectively repress epithelial markers such as E-cadherin, Zona occludens-1, claudins, and cytokeratin while activating mesenchymal markers including vimentin, N-cadherin, fibronectin, and α-smooth muscle actin [45] [46]. These transcriptional changes drive fundamental cellular rearrangements including loss of apical-basal polarity, dissolution of tight junctions and adherent junctions, and reorganization of the actin cytoskeleton to form protrusive structures like filopodia and lamellipodia that enable cell migration [45] [46].
Several principal signaling pathways regulate EMT through activation of these transcription factors. The TGF-β pathway activates SMAD proteins (SMAD2/3/4 complex) that translocate to the nucleus to repress epithelial genes and activate mesenchymal genes [46]. Wnt/β-catenin signaling stabilizes β-catenin, which partners with LEF/TCF transcription factors to promote EMT gene expression programs [45]. Additional pathways including Notch, ERK, and hypoxia-induced factors further modulate EMT progression through context-specific mechanisms [45]. During gastrulation, these pathways are activated by morphogen gradients along the anterior-posterior axis, with high Wnt/β-catenin, TGF-β, and fibroblast growth factor signaling driving the EMT program in posterior epiblast cells [26].
Figure 1: Core Signaling Pathways Regulating EMT. Multiple extracellular signaling pathways converge on key transcription factors that coordinately repress epithelial markers and activate mesenchymal markers, driving cytoskeletal reorganization and cellular transformation.
EMT in Primitive Streak Formation
During mammalian gastrulation, the primitive streak forms as a transient structure through which epiblast cells ingress to form mesoderm and definitive endoderm. Recent single-cell studies combining lineage tracing with transcriptomic analysis have revealed that rather than a uniform EMT program, different germ layers employ distinct morphogenetic strategies [26]. Mesoderm formation follows classical EMT with strong upregulation of Snail1, complete E- to N-cadherin switching, and acquisition of mesenchymal morphology enabling cell migration. In contrast, definitive endoderm formation occurs through a Snail1-independent partial EMT program characterized by maintenance of E-cadherin expression with coordinated upregulation of N-cadherin, allowing cells to ingress while retaining epithelial characteristics [26].
This divergence in EMT programs is regulated by key transcription factors: Brachyury (T) drives mesodermal EMT while Foxa2 acts as an epithelial gatekeeper and EMT suppressor to shield endoderm progenitors from complete mesenchymal transition [26]. Foxa2+ endoderm progenitors ingress distally to the anatomical primitive streak, transitioning from the epiblast while maintaining epithelial features, then intercalate into the visceral endoderm layer without undergoing a subsequent MET, challenging the long-standing EMT-MET cycle paradigm for endoderm formation [26].
Transcriptomic Approaches and Methodologies
Single-Cell RNA Sequencing (scRNA-seq)
Single-cell transcriptomics has revolutionized the study of EMT in primitive streak formation by enabling the resolution of heterogeneous cell states and transitional populations along developmental trajectories. The standard workflow begins with cell isolation and encapsulation, followed by reverse transcription, cDNA amplification, library preparation, and sequencing [47] [48] [26]. For embryonic studies, fluorescent reporter knock-in systems (e.g., Foxa2-Venus, Sox17-mCherry, T-GFP) enable fluorescence-activated cell sorting (FACS) to isolate specific progenitor populations prior to scRNA-seq, dramatically enhancing resolution of rare transitional states [26].
Key Analytical Frameworks:
- Trajectory Inference: Pseudotime algorithms (Monocle, Slingshot) reconstruct developmental trajectories from scRNA-seq data, ordering cells along progression paths from epithelial to mesenchymal states [48].
- RNA Velocity: ScVelo and CellRank analyze unspliced/spliced mRNA ratios to infer directional state transitions and fate probabilities, revealing lineage driver genes during mesoderm and endoderm segregation [26].
- Differential Expression Analysis: Identifies genes significantly upregulated in specific states (e.g., EMT intermediate states) using statistical frameworks like DESeq2 [47].
A recent multi-cancer scRNA-seq study analyzing EMT trajectories across twelve datasets identified 32 genes consistently upregulated in intermediate EMT states, most encoding extracellular or plasma membrane proteins including ITGB4, ITGA6, SFN, SNCG, and WNT9A [48]. These intermediate state markers were largely distinct from canonical EMT genes, highlighting the unique transcriptional identity of hybrid E/M states.
Bulk RNA Sequencing
While scRNA-seq provides superior resolution of cellular heterogeneity, bulk RNA sequencing remains valuable for profiling population-level transcriptomic changes during EMT. Applications include:
- Time-series analysis of EMT progression in response to developmental signals (TGF-β, Wnt, FGF) [45]
- Comparative analysis of epithelial, intermediate, and mesenchymal states from sorted populations [26]
- Integration with proteomic data to identify post-transcriptional regulatory mechanisms [49]
Bulk RNA sequencing typically provides higher sequencing depth per sample, enabling detection of low-abundance transcripts, though it obscures cellular heterogeneity within samples. Experimental design should include appropriate biological replicates (minimum n=3), randomization, and batch effect correction to ensure robust results.
Proteomic and Phosphoproteomic Approaches
Mass Spectrometry-Based Proteomics
Proteomic approaches provide essential insights into EMT mechanisms by quantifying protein expression changes, post-translational modifications, and protein-protein interactions that cannot be inferred from transcriptomic data alone. Several mass spectrometry strategies are employed in EMT research:
Tandem Mass Tag (TMT) Proteomics:
- Principle: Multiplexed isobaric labeling enabling simultaneous quantification of multiple samples [49]
- Workflow: Protein extraction and digestion → TMT labeling → basic reversed-phase LC fractionation → high-resolution tandem MS → database searching → quantification
- Application in EMT: Temporal profiling of protein expression changes during RPE-EMT identified 532 differentially regulated proteins, revealing both commonalities and distinctions with malignancy-associated EMT [49]
Data-Independent Acquisition (DIA):
- Principle: MS2-level quantification without precursor isolation, improving reproducibility and quantitative accuracy [49]
- Application: Validation of TMT findings, particularly for low-abundance proteins and post-translational modifications
Experimental Design Considerations:
- Include minimum three biological replicates per condition
- Implement blocking designs to account for technical variability
- Use spike-in standards for normalization when comparing different cellular states
- Combine with transcriptomic data to identify post-transcriptional regulation
Phosphoproteomics in EMT Signaling
Phosphoproteomic analysis provides critical insights into signaling network dynamics during EMT. The BAG5 study employed phosphoproteomics to identify phosphorylation changes associated with cytoskeletal remodeling and metabolic reprogramming in non-small cell lung cancer [50]. Standard protocols involve:
- Protein extraction under denaturing conditions with phosphatase inhibitors
- Trypsin digestion followed by phosphopeptide enrichment (TiO2, IMAC, or antibody-based)
- LC-MS/MS analysis with collision-induced dissociation or electron-transfer dissociation
- Phosphosite localization using algorithms like AScore or PTM-RS
- Pathway analysis to identify activated kinase networks
Integration of phosphoproteomic data with transcriptomic and proteomic datasets enables comprehensive reconstruction of signaling networks driving EMT progression and identifies potential therapeutic targets for modulating these pathways.
Multi-Omic Integration Strategies
Computational Integration Frameworks
The integration of multiple omics datasets presents significant computational challenges due to data heterogeneity, technical noise, and missing values. Several computational frameworks have been developed specifically for multi-omics integration in biological research:
Table 1: Multi-Omics Integration Methods
| Method | Approach | Strengths | Limitations |
|---|---|---|---|
| MOSA [51] | Unsupervised deep learning (variational autoencoder) | Handles data sparsity, generates synthetic data, explains features via SHAP | Requires substantial computational resources |
| MOFA+ [51] | Factor analysis (linear dimensionality reduction) | Interpretable factors, handles missing data, multiple data types | Assumes linear relationships between omics layers |
| MixOmics [51] | Multivariate analysis (projection to latent structures) | Provides variable selection, multiple integration modes | Limited to paired samples across omics |
| WGCNA [47] | Weighted gene co-expression network analysis | Identifies co-expression modules, relates modules to traits | Primarily for single-omic analysis followed by integration |
Deep Learning for Multi-Omic Data Augmentation
The MOSA (Multi-Omic Synthetic Augmentation) framework represents a significant advancement in multi-omics integration, addressing the critical challenge of data sparsity in large-scale cancer cell line datasets [51]. This unsupervised deep learning model employs a variational autoencoder architecture with several innovative features:
- Late Integration Approach: Separate encoders for each omic type with concatenated latent embeddings
- Conditional Integration: Incorporation of genetic background as conditional variables
- Asymmetrical Design: Uses most variable features as encoder input while reconstructing all features
- Whole-Omic Dropout: Regularization technique masking complete omic layers during training
When applied to the Cancer Dependency Map (DepMap) consortium data spanning seven omic types across 1,523 cancer cell lines, MOSA increased complete multi-omic profiles by 32.7%, significantly enhancing statistical power for identifying genetic associations and clustering relationships [51]. The model successfully reconstructed independent drug response datasets (Pearson's r=0.87), outperforming other integration methods and demonstrating robust generalizability [51].
Figure 2: Multi-Omic Integration Workflow Using Deep Learning. The MOSA framework processes multiple omics data types through separate encoders, integrates them in a joint latent space conditioned on genetic background, and reconstructs complete omics profiles, enabling data augmentation and imputation of missing values.
Experimental Protocols for Key Methodologies
Time-Resolved Lineage Tracing with scRNA-seq
This protocol combines fluorescent reporter systems with single-cell transcriptomics to resolve lineage relationships and transcriptional dynamics during EMT in gastrulation [26].
Materials:
- Foxa2-Venus fusion (FVF) and/or Sox17-mCherry fusion (SCF) knock-in reporter mouse embryonic stem cells
- Embryo culture media and differentiation conditions
- Fluorescence-activated cell sorter (FACS) with appropriate lasers and detectors
- Single-cell RNA sequencing platform (10X Genomics, Smart-seq2, etc.)
- Bioinformatics tools: Seurat, ScVelo, CellRank, Monocle
Procedure:
- Differentiation: Differentiate reporter mESCs toward mesendodermal lineages using established protocols with activin A, Wnt3a, and BMP4 signaling modulation [26].
- Time-Series Sampling: Collect cells at multiple time points (e.g., days 0, 1, 2, 3, 4 of differentiation) to capture transitional states.
- FACS Isolation: Dissociate cells to single-cell suspension and sort based on reporter fluorescence intensity into distinct populations (e.g., FVFlow epiblast progenitors, FVFhigh transitory progenitors, FVFhigh/Sox17+ definitive endoderm) [26].
- scRNA-seq Library Preparation: Process sorted cells using appropriate scRNA-seq technology (10X Genomics for high-throughput, Smart-seq2 for greater depth).
- Sequencing: Sequence libraries following platform-specific recommendations (minimum 50,000 reads per cell for 10X Genomics).
- Computational Analysis:
- Quality control and filtering using Seurat (remove cells with <200 genes or >10% mitochondrial reads)
- Normalization and integration of multiple time points
- Clustering and cell type annotation using marker genes
- Trajectory inference using pseudotime algorithms
- RNA velocity analysis to infer directional state transitions
- Differential expression analysis across transitions
Integrative Multi-Omic Analysis of EMT
This protocol outlines a comprehensive approach for integrating transcriptomic, proteomic, and phosphoproteomic data to identify coordinated molecular events during EMT [50] [49].
Materials:
- Cell lines or primary cells undergoing EMT (TGF-β treatment, genetic manipulation, etc.)
- RNA extraction kit (e.g., Qiagen RNeasy)
- Protein extraction buffer (e.g., 8M urea, 100mM Tris-HCl pH 8.0) with protease and phosphatase inhibitors
- Tandem mass tag (TMT) reagents (Thermo Scientific)
- LC-MS/MS system with nanoflow liquid chromatography and high-resolution mass spectrometer
- Bioinformatics tools: MaxQuant, MOSA, MOFA+, Ingenuity Pathway Analysis
Procedure:
- Experimental Design:
- Establish EMT model system with appropriate controls and replicates (minimum n=3 biological replicates)
- Collect samples at multiple time points to capture dynamic changes
- Process aliquots of the same samples for different omics analyses
Transcriptomic Analysis:
- Extract total RNA following manufacturer's protocol
- Assess RNA quality (RIN >8.0 required)
- Prepare RNA-seq libraries using polyA selection or ribosomal RNA depletion
- Sequence to minimum depth of 30 million reads per sample
- Process data: quality control (FastQC), alignment (STAR), quantification (featureCounts), differential expression (DESeq2)
Proteomic Analysis:
- Extract proteins in denaturing buffer
- Digest proteins using trypsin/Lys-C mix
- Label peptides with TMT reagents following manufacturer's protocol
- Fractionate using basic reversed-phase chromatography
- Analyze by LC-MS/MS with data-dependent acquisition
- Process data: database search (MaxQuant), normalization, differential abundance (Limma)
Phosphoproteomic Analysis:
- Enrich phosphopeptides from protein digest using TiO2 or IMAC
- Analyze by LC-MS/MS with optimized methods for phosphopeptide detection
- Process data: phosphosite localization, differential phosphorylation
Multi-Omic Integration:
- Preprocess datasets to common gene/protein identifiers
- Apply integration method (MOSA, MOFA+) to identify shared and unique variation across omics layers
- Perform pathway enrichment analysis on integrated features
- Validate key findings using orthogonal methods (Western blot, immunofluorescence, functional assays)
The Scientist's Toolkit: Essential Research Reagents
Table 2: Key Research Reagents for EMT and Multi-Omic Studies
| Reagent Category | Specific Examples | Application/Function | Considerations |
|---|---|---|---|
| EMT Inducers | TGF-β1-3, Wnt3a, FGF2, TNF-α, HIF-1α stabilizers | Activate EMT signaling pathways in model systems | Concentration and timing critically affect EMT progression and state heterogeneity |
| Lineage Reporters | Foxa2-Venus, Sox17-mCherry, T-GFP knock-in cells [26] | Fate mapping and isolation of specific progenitor populations | Requires validation of reporter fidelity and minimal perturbation of endogenous function |
| EMT Marker Antibodies | E-cadherin, N-cadherin, vimentin, ZO-1, Snail, Slug, Twist | Immunofluorescence, Western blot, flow cytometry validation of EMT states | Context-dependent expression patterns require multiple marker validation |
| Single-Cell Platforms | 10X Genomics Chromium, BD Rhapsody, Smart-seq2 | High-throughput scRNA-seq of EMT populations | Platform choice balances cell throughput, sequencing depth, and cost |
| Mass Spectrometry Reagents | TMTpro 16-plex, iRT peptides, TiO2/IMAC phospho-enrichment | Multiplexed proteomic and phosphoproteomic quantification | Consider plex level needed for experimental design and normalization strategies |
| Bioinformatics Tools | Seurat, Scanpy, MOSA, MOFA+, ScVelo, CellRank | Analysis and integration of multi-omics datasets | Computational resource requirements and learning curve vary significantly |
Molecular profiling through transcriptomic, proteomic, and multi-omic approaches has fundamentally transformed our understanding of EMT in primitive streak formation and cancer progression. The integration of these complementary data types, particularly through advanced computational frameworks like MOSA, has revealed unprecedented details about the regulatory networks, intermediate states, and dynamic transitions that characterize these processes. The finding that distinct germ layers employ different morphogenetic programs—classical EMT for mesoderm versus partial EMT for definitive endoderm—exemplifies how multi-omic approaches can challenge and refine long-standing biological paradigms [26].
Future directions in this field will likely focus on several key areas: First, spatial multi-omics technologies will enable the mapping of molecular events within their native tissue context, revealing how positional information influences EMT progression. Second, the integration of single-cell epigenomic data (ATAC-seq, ChIP-seq) with transcriptomic and proteomic datasets will provide more comprehensive views of regulatory mechanisms. Third, improved dynamical modeling approaches will enhance our ability to predict EMT trajectories and outcomes from initial molecular states. Finally, the translation of these fundamental insights into therapeutic applications, particularly in targeting EMT-related drug resistance in cancer, represents a critical frontier. As these technologies continue to evolve and become more accessible, they will undoubtedly yield further transformative insights into the molecular choreography of epithelial-mesenchymal transitions in development and disease.
Live Imaging and Quantification of Cell Ingression and Migration
Epithelial-to-mesenchymal transition (EMT) is a fundamental process in development, cancer metastasis, and fibrosis, during which epithelial cells lose cell-cell adhesion and acquire migratory mesenchymal properties [52] [14]. In mammalian development, the primitive streak serves as the principal site for gastrulation EMT, where epiblast cells ingress to form the mesoderm and endoderm [6]. Understanding the cellular dynamics of this process requires sophisticated live imaging approaches that can capture and quantify cell behaviors in real time. This technical guide outlines current methodologies for visualizing and analyzing cell ingression and migration, with specific focus on applications within primitive streak research.
Live Imaging Modalities for EMT Analysis
Selecting the appropriate imaging modality depends on the experimental model, required resolution, and duration of imaging. The following table compares key live imaging approaches relevant to studying EMT in primitive streak contexts:
Table 1: Live Imaging Modalities for EMT and Cell Migration Studies
| Imaging Modality | Spatial Resolution | Temporal Capability | Primary Applications | Key Advantages | Model Systems |
|---|---|---|---|---|---|
| Confocal Microscopy | High (subcellular) | Hours to days [53] | Mouse gastrulation EMT; cell tracking in 3D [6] [53] | Optical sectioning; robust commercial systems | Mouse embryos, crustacean limbs [6] [53] |
| Two-Photon Microscopy | High (tissue penetration) | Hours to days [39] | Intravital tumor imaging; deep tissue visualization | Reduced photodamage; deeper tissue penetration | Mammary tumors in mouse models [39] |
| Spinning Disk Confocal | High | Hours to days | Rapid 4D imaging of dynamic processes | Faster imaging with reduced photobleaching | Developing embryos [52] |
| Light-Sheet Microscopy | Medium to High | Days [6] | Long-term developmental processes; large samples | Extremely low phototoxicity; fast imaging | Whole mouse embryos [6] |
| Incucyte Live-Cell Analysis | Medium | Days [54] | Scratch wound migration assays; high-throughput screening | Automated, reproducible; integrated analysis | Cell monolayers in vitro [54] |
Experimental Models and Preparation
Mouse Primitive Streak Models
The mouse embryo remains the primary model for studying gastrulation EMT in mammals. Key technical considerations include:
- Genetic Reporters: Utilizing ZO-1-GFP fusion proteins to visualize tight junctions and apical surfaces of epiblast cells, or membrane-localized reporters (Rosa26mT/mG) to track entire plasma membranes during ingression [6].
- Ex Utero Culture: Embryos need to be kept intact without microdissection to maintain epiblast integrity during time-lapse imaging [6].
- Imaging Challenges: The epiblast's cup-shaped curvature and deep location (up to 60 µm from the objective) require imaging through adjacent tissue layers [6].
Recent studies using these approaches have revealed that epiblast cells constrict their apical surfaces in a "ratchet-like pulsed fashion" through asynchronous shrinkage of apical cell-cell junctions during ingression [6].
In Vitro Migration Assays
For simplified systems, scratch wound assays provide quantitative data on cell migration:
- Incucyte Scratch Wound Assay: Utilizes a 96-Well WoundMaker tool to create precise, uniform wounds in cell monolayers cultured in Imagelock plates [54].
- Protocol: Cells are seeded into specialized plates, incubated overnight, wounded after reaching confluence, then treated with experimental media. The integrated software automatically detects and registers wound locations for consistent analysis [54].
- Quantification: Migration is typically measured using Relative Wound Density (RWD), which compares cell density inside the wound to the surrounding area over time [54].
Quantitative Analysis of Cell Dynamics
Metrics for Cell Ingression
In mouse primitive streak studies, quantitative analysis reveals specific ingression dynamics:
- Ingression Timing: 44 ± 2% of cells within the primitive streak domain constrict and ingress within a 1-hour period (mean ± s.e.m., n=378 cells, three embryos) [6].
- Ingression Patterns: 48% of cells ingress as isolated events (>30 min apart from neighbors), while 52% ingress as pairs or small groups (<30 min apart) [6].
- Apical Constriction Dynamics: Cells undergo pulsed apical constriction through asynchronous shrinkage of apical junctions, which can be quantified by measuring apical surface area over time [6].
Metrics for Cell Migration
In scratch wound and other migration assays, key quantitative parameters include:
- Relative Wound Density (RWD): A primary metric that calculates the density of cells in the wound area relative to the density outside the wound area [54].
- Migration Rate: The speed of wound closure over time, often measured in µm/hour or % closure/hour.
- Pharmacological Response: IC50 values for inhibitory compounds can be determined from concentration-response curves [54].
Table 2: Quantitative Metrics for Cell Ingression and Migration Analysis
| Metric | Definition | Application Context | Typical Values/Examples |
|---|---|---|---|
| Relative Wound Density (RWD) | Ratio of cell density inside vs. outside wound | Scratch wound assays [54] | HeLa WT: 60% at 48h; PI3K KO: 40% at 48h [54] |
| Apical Surface Area | Area of apical cell surface facing lumen | Primitive streak ingression [6] | Progressive reduction during pulsed constriction |
| Ingression Percentage | Proportion of cells ingressing within time window | Primitive streak analysis [6] | 44 ± 2% per hour in mouse primitive streak |
| IC50 Migration | Compound concentration yielding 50% migration inhibition | Pharmacological assessment [54] | Cell-type dependent (e.g., Cytochalasin D in HT-1080 vs. MDA-MB-231) |
| Coordinated Ingression | Cells ingressing within 30 minutes of neighbors | Collective behaviors in EMT [6] | 52% of cells ingress as pairs or groups |
Signaling Pathways in Primitive Streak EMT
The following diagram illustrates key signaling pathways regulating EMT and cell ingression in the primitive streak context:
Signaling Pathways in Primitive Streak EMT
Experimental Protocols
Mouse Primitive Streak Live Imaging Protocol
This protocol enables visualization of EMT in mouse gastrulation embryos [6]:
- Embryo Preparation: Collect mouse embryos at E6.5-E7.5 days post-coitum in pre-warmed DMEM/F-12 medium.
- Genetic Labeling: Use ZO-1-GFP transgenic reporters to label tight junctions and apical surfaces.
- Mounting: Position embryos in glass-bottom dishes with the primitive streak oriented toward the coverslip.
- Culture Conditions: Maintain embryos in rat serum at 37°C with 5% CO2 during imaging.
- Image Acquisition: Acquire z-stacks every 5-10 minutes for 4-12 hours using confocal or two-photon microscopy.
- Data Analysis: Segment cell membranes and track apical surface area changes, ingression timing, and cell trajectories.
Scratch Wound Migration Assay Protocol
This standardized protocol enables quantitative analysis of 2D cell migration [54]:
- Cell Seeding: Seed cells into Incucyte Imagelock 96-well plates at optimal density (e.g., 30,000-50,000 cells/well).
- Incubation: Incubate overnight at 37°C with 5% CO2 to form confluent monolayers.
- Wound Creation: Use the Incucyte WoundMaker tool to create uniform scratches in all wells.
- Washing and Treatment: Wash away dislodged cells and add treatment media at 100 µL per well.
- Image Acquisition: Place plate in Incucyte Live-Cell Analysis System; acquire phase-contrast images every 2 hours for 24-72 hours.
- Quantitative Analysis: Use Incucyte Scratch Wound Analysis Software Module to calculate RWD metrics automatically.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Live Imaging of Cell Ingression and Migration
| Reagent/ Tool | Function | Example Application | References |
|---|---|---|---|
| Incucyte WoundMaker | Creates uniform scratches in cell monolayers | Standardized wound healing assays | [54] |
| ZO-1-GFP Reporter | Labels tight junctions and apical surfaces | Visualizing apical constriction in mouse epiblast | [6] |
| Rosa26mT/mG Reporter | Membrane-localized fluorescent reporter | Tracking cell membranes during ingression | [6] |
| Fsp1-Cre; Rosa26-RFP-GFP System | EMT-driven fluorescent color switching | Tracking EMT in breast cancer models | [39] |
| Mitomycin C | Anti-proliferative agent | Distinguishing migration from proliferation | [54] |
| Cytochalasin D | Actin polymerization inhibitor | Pharmacological inhibition of migration | [54] |
| TGF-β | EMT-inducing growth factor | Inducing EMT in A549/Vim-RFP cells | [55] |
Data Analysis and Computational Approaches
Single-Cell Trajectory Analysis
Advanced computational methods enable quantitative analysis of EMT dynamics:
- Morphology/Texture Feature Space: Cell states can be represented in a multi-dimensional space combining cell body shape (296 degrees of freedom) and vimentin texture features (13 Haralick features) [55].
- Reaction Coordinate Reconstruction: From single-cell trajectories, researchers can reconstruct quasi-potentials to understand transition dynamics between epithelial and mesenchymal states [55].
- Bifurcation Analysis: EMT may proceed through saddle-node bifurcations where the original stable epithelial attractor collides with saddle points before relaxing to a new mesenchymal state [55].
Cell Tracking and Lineage Analysis
In regenerating systems and developing embryos:
- Long-Term Tracking: Methods have been developed to track individual cells over up to 10 days at 20-minute intervals in regenerating crustacean limbs [53].
- Computer-Assisted Cell Tracking: Combined with appropriate cell-type-specific markers, these approaches can determine complete cell lineages for regenerated tissues [53].
- Fixed/Track Correlation: After live imaging, tissues can be fixed and stained to identify cell fates, correlating dynamic behaviors with final differentiation states [53].
Technical Challenges and Optimization Strategies
Minimizing Photodamage
Long-term live imaging requires balancing resolution with cell viability:
- Wavelength Selection: Imaging at long wavelengths (e.g., using H2B-mRFPruby) reduces photodamage during extended time-lapse experiments [53].
- Temporal Resolution Optimization: For processes like leg regeneration, 20-minute intervals sufficiently capture cell divisions and movements while minimizing light exposure [53].
- Light-Sheet Considerations: While ideal for reducing phototoxicity, current light-sheet designs may be incompatible with imaging through glass coverslips required for certain immobilized specimens [53].
Distinguishing Migration from Proliferation
In scratch wound assays, careful experimental design can separate these processes:
- Anti-Proliferative Agents: Mitomycin C (MMC) pre-treatment (e.g., 50µM for 4 hours) helps distinguish migration-driven from proliferation-driven wound closure [54].
- Cell-Type Specificity: The contribution of proliferation varies by cell type; MMC has no measurable impact on migration in MDA-MB-231 cells, but reduces apparent migration in BxPC3 cells where proliferation contributes significantly to closure [54].
The following diagram illustrates a optimized experimental workflow for primitive streak live imaging:
Primitive Streak Live Imaging Workflow
Epithelial-mesenchymal transition (EMT) is a fundamental cellular process during which epithelial cells lose their polarity and cell-cell adhesion and acquire a migratory, mesenchymal phenotype [9] [13]. In the context of early embryogenesis, particularly during primitive streak formation and gastrulation, a specific subtype known as Type 1 EMT occurs [13]. This process is essential for the generation of the mesoderm and endoderm from the epiblast layer, forming the primary mesenchyme that gives rise to diverse tissues and organs [13]. The functional validation of genes and signaling pathways orchestrating this critical developmental event is paramount to understanding congenital disorders and the underpinnings of cellular plasticity, which is re-activated in pathological conditions such as cancer and fibrosis [13] [56].
Modern functional genomics aims to move beyond mere associations and establish causal relationships between genes and biological phenotypes [57]. Loss-of-function strategies, which involve disrupting a gene and observing the resulting phenotypic consequences, are a cornerstone of this approach [57]. The advent of the CRISPR-Cas9 system has revolutionized loss-of-function studies, providing a precise, scalable, and efficient method for targeted gene disruption [58] [57]. This technical guide details the application of CRISPR-Cas9 and related methodologies for the functional validation of genetic regulators within the specific context of EMT in primitive streak research.
Core Principles of CRISPR-Cas9 Mediated Loss-of-Function
The CRISPR-Cas9 system comprises two core components: the Cas9 nuclease, which creates double-strand breaks in DNA, and a guide RNA (gRNA), which directs Cas9 to a specific genomic locus complementary to its sequence [57]. The cell's repair of these breaks via the error-prone non-homologous end joining (NHEJ) pathway often results in small insertions or deletions (indels). When these indels occur within a protein-coding exon, they can cause a frameshift mutation, leading to a premature stop codon and a truncated, non-functional protein [57].
A significant advancement in this field is the "perturbomics" approach, which leverages CRISPR-Cas screens for the systematic, large-scale functional annotation of genes based on the phenotypic changes induced by their perturbation [57]. This is particularly powerful for moving from observational data in EMT, such as gene expression signatures from single-cell RNA sequencing, to causal validation [57] [56].
Table 1: Key CRISPR-Cas9 System Components for Loss-of-Function Studies
| Component | Description | Function in Loss-of-Validation |
|---|---|---|
| Cas9 Nuclease | Enzyme from bacterial immune system that cuts DNA. | Induces double-strand breaks at targeted genomic locations. |
| Guide RNA (gRNA) | A short RNA sequence that combines tractRNA and crRNA. | Specifies the DNA target site via complementary base pairing. |
| Repair Template | (For knock-in) Donor DNA template for HDR. | Not typically used in standard knockout strategies; used for precise edits. |
| Delivery Vector | Viral (e.g., Lentivirus, Adenovirus) or non-viral vehicle. | Introduces Cas9 and gRNA constructs into target cells. |
Experimental Design and Workflow for EMT Studies
A typical workflow for CRISPR-Cas9 mediated functional validation in a primitive streak or EMT model involves several critical steps, from target identification to phenotypic analysis.
Target Selection and gRNA Design
The process begins with the selection of candidate genes, which may originate from genomic studies of the primitive streak or transcriptomic analyses of cells undergoing EMT [58] [59]. For each target gene, multiple gRNAs (typically 3-5) are designed to target early exons of the coding sequence to maximize the likelihood of a disruptive frameshift. gRNA design tools (e.g., from the Broad Institute or Benchling) are used to minimize off-target effects by ensuring the chosen gRNA sequences have minimal similarity to other genomic regions [60].
Delivery of CRISPR Components
Effective delivery of the Cas9 nuclease and gRNA into the target cells is crucial. For in vitro studies in 2D cell culture or 3D organoids, lentiviral or adenoviral vectors are commonly used due to their high efficiency [58]. The adenoviral vector Ad5-USEC, for instance, has been successfully employed to co-express a gRNA and Cre recombinase for cell-type-specific targeting in complex models [58]. For research requiring high physiological relevance, organoid systems provide an excellent platform that more closely mimics the in vivo environment than traditional 2D cultures [61] [57].
Validation of Gene Knockout
Following delivery, the success of gene editing must be confirmed. This is typically a two-tiered process:
- Genotypic Validation: Genomic DNA from targeted cells is amplified by PCR around the gRNA target site and subjected to Sanger sequencing or next-generation sequencing to detect the spectrum of indels and calculate the editing efficiency [58].
- Protein Validation: Western blot analysis is used to confirm the reduction or absence of the target protein, which is the ultimate proof of a successful knockout [58].
Signaling Pathways in Primitive Streak EMT and Targeting Strategies
The EMT occurring during primitive streak formation is orchestrated by a conserved set of signaling pathways and transcription factors. Understanding these pathways provides the context for selecting targets for functional validation [13] [62].
The canonical Wnt signaling pathway is a primary driver, with Wnt3 being essential for the initiation of gastrulation [13]. This is closely coordinated with signaling from the TGF-β superfamily, including Nodal and Vg1, and with FGF receptor signaling [13]. These pathways converge to activate key EMT-transcription factors (EMT-TFs) such as Snail, Eomes, and Mesps, which pleiotropically repress epithelial genes (e.g., E-cadherin, occludins) and activate the mesenchymal program [9] [13]. The neural crest, another classic model of developmental EMT, utilizes similar pathways, including BMP, Wnt, and FGF signaling, to initiate delamination and migration [62].
Table 2: Key Signaling Pathways and Factors in Primitive Streak EMT
| Pathway/Factor | Role in Primitive Streak EMT | Potential CRISPR Target |
|---|---|---|
| Wnt Signaling | Master regulator; initiates gastrulation EMT. | Wnt3, β-catenin (Ctnnb1) |
| TGF-β/Nodal | Cooperates with Wnt; induces mesoderm formation. | Nodal, Tgfb1, Smad2/3 |
| FGF Signaling | Cooperates with Wnt; regulates EMT ingression. | Fgf receptors, downstream kinases |
| Transcription Factors | Execute EMT program; repress E-cadherin. | Snail (Snai1), Eomes, Mesp1/2 |
| Cell Adhesion | Target of EMT-TFs; loss enables cell migration. | Cdh1 (E-cadherin), Cldns |
Advanced CRISPR Methodologies and Applications
Beyond simple knockout, the CRISPR toolkit has expanded to include more sophisticated perturbation methods that are highly relevant for dissecting the nuances of EMT.
CRISPR Screening in EMT Models
CRISPR knockout screens can be performed genome-wide or on a focused gene set to unbiasedly identify all genetic factors that confer a specific phenotype related to EMT [57] [59]. For example, a parallel genome-wide CRISPR screen comparing epithelial-like and mesenchymal-like pancreatic cancer cells identified distinct genetic determinants of immune evasion, such as Egfr and Mfge8, which were specifically required for the resistance of mesenchymal cells to T-cell killing [59]. This approach can be applied to primitive streak models to screen for genes essential for mesoderm formation or migration.
Beyond Knockout: CRISPRi and CRISPRa
Nuclease-inactive dCas9 can be fused to repressive (e.g., KRAB) or activating (e.g., VPR) domains to create CRISPR interference (CRISPRi) or CRISPR activation (CRISPRa) systems, respectively [57]. These tools allow for reversible, tunable gene knockdown or overexpression without altering the underlying DNA sequence. This is particularly useful for validating the function of essential genes where a knockout would be lethal, or for studying the effects of overexpressing specific EMT-TFs in a controlled manner. CRISPRi also enables the targeting of non-coding genomic elements, such as enhancers that regulate EMT genes [57].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Research Reagent Solutions for CRISPR-based Functional Validation
| Reagent / Tool | Function | Example Application in EMT |
|---|---|---|
| CRISPR-Cas9 Plasmid Libraries | Pooled collection of gRNA expression vectors. | Genome-wide screens for EMT regulators [59]. |
| Lentiviral/Adenoviral Vectors | High-efficiency delivery of CRISPR constructs. | Transduction of primary cells or organoids [58]. |
| Cre-Activatable Cas9 Alleles | Enables cell-type-specific gene editing. | Spatial-temporal knockout in specific embryonic lineages [58]. |
| dCas9-KRAB / dCas9-VPR | Targeted gene repression (CRISPRi) or activation (CRISPRa). | Studying essential genes or overexpressing EMT-TFs [57]. |
| Single-Cell RNA-seq Kits | Transcriptomic profiling of individual cells. | Characterizing heterogeneous EMT states after perturbation [56]. |
| Lineage Tracing Models | Tracks the fate of specific cell populations in vivo. | Validating the contribution of knocked-out cells to mesodermal lineages. |
CRISPR-Cas9 mediated loss-of-function strategies provide a powerful and flexible framework for the functional validation of genes controlling epithelial-mesenchymal transition. When applied to the context of primitive streak research, these methods allow researchers to move from correlative observations to causal understanding, systematically dissecting the genetic hierarchy that orchestrates one of the most critical events in embryonic development. The continuous development of more precise editing tools and complex in vitro models, such as organoids, promises to further enhance the resolution and physiological relevance of these functional studies, deepening our grasp of both development and disease.
Resolving Complexities: EMT Plasticity, Partial States, and Technical Challenges
Epithelial-mesenchymal transition (EMT) is a fundamental cellular process through which epithelial (E) cells lose their adherens junctions and apicobasal polarity, acquiring migratory and invasive mesenchymal (M) characteristics. Historically conceptualized as a binary process, our understanding has evolved to recognize EMT as a dynamic spectrum of intermediate states. Hybrid epithelial/mesenchymal (E/M) states represent a critical intermediate phase where cells co-express both epithelial and mesenchymal markers, exhibiting a unique cellular plasticity that enhances their tumorigenic potential [63]. This plastic transient state enables adaptation to microenvironmental cues and drives progression in multiple pathological contexts, particularly cancer metastasis and therapeutic resistance [63] [64].
The study of EMT finds its roots in developmental biology, with the primitive streak serving as a foundational model. During gastrulation in amniotes, a subset of cells from the epiblast moves to the midline to form the primitive streak, undergoing EMT to internalize and generate mesoderm and endoderm [65]. These early embryonic cells demonstrate remarkable plasticity, operating without fully mature junctional complexes and resulting in mesenchymal-like cells that don't necessarily exhibit a full-blown mesenchymal phenotype [4]. This embryonic plasticity is re-activated in pathological contexts, making understanding the primitive streak essential for deciphering the mechanisms governing hybrid E/M states in cancer.
Defining Hybrid E/M States: Beyond the Binary Paradigm
Conceptual Framework and Cellular Characteristics
Hybrid E/M phenotypes represent a departure from the traditional binary view of EMT. Cells in these intermediate states lose apical-basal polarity but retain some cell-cell adhesions, gaining motility while maintaining limited connections to neighboring cells [63]. This enables collective cell migration, where cells migrate as cohesive groups rather than individually, balancing migratory capability with protective cell-cell signaling [63]. The hybrid E/M state is not a single defined point but encompasses a spectrum of intermediate states with varying ratios of epithelial to mesenchymal characteristics, creating dynamic cellular ecosystems within tumors that facilitate invasion, metastasis, and treatment resistance [4].
The plasticity of hybrid E/M states represents their defining feature. Cells can dynamically shift their position along the E/M spectrum in response to microenvironmental signals, with the hybrid E/M subpopulation demonstrating the greatest plasticity, capable of giving rise to both E and M phenotypes [63]. This plasticity enables adaptation to diverse challenges during metastatic progression, from detachment from the primary tumor to survival in circulation and colonization of distant sites.
Molecular Markers and Regulatory Networks
The identification of hybrid E/M states relies on monitoring specific molecular markers that signify the simultaneous presence of epithelial and mesenchymal characteristics.
Table 1: Key Molecular Markers for Identifying Hybrid E/M States
| Marker Type | Specific Markers | Expression in Hybrid E/M States | Functional Significance |
|---|---|---|---|
| Epithelial Markers | E-cadherin (CDH1), Cytokeratins, Occludin | Reduced but not absent | Maintains limited cell-cell adhesion |
| Mesenchymal Markers | N-cadherin (CDH2), Vimentin, Fibronectin | Elevated but not maximal | Enhances motility and matrix interaction |
| Hybrid State Indicators | P-cadherin, Slug, Integrin-β4 (ITGB4) | Specifically elevated | Facilitates collective migration |
| Surface Markers | CD104+CD44hi, CD24hi/CD44hi | Characteristic expression patterns | Enables identification and isolation |
The regulatory network governing hybrid E/M states involves sophisticated transcriptional and post-transcriptional control. Core EMT transcription factors (EMT-TFs)—including SNAI1, SNAI2 (Slug), ZEB1, ZEB2, and TWIST—operate in interconnected feedback loops that stabilize intermediate states [4] [9]. Phenotypic stability factors (PSFs) such as GRHL2, OVOL2, and NUMB help maintain the hybrid E/M state by preventing complete transition to either terminal state [63] [64]. These regulators respond to diverse signaling inputs from the tumor microenvironment, including TGF-β, Wnt, and inflammatory signals, allowing dynamic adjustment of the cellular phenotype along the E/M spectrum [4] [66].
Quantitative Assessment of Hybrid E/M States
Transcriptomic and Proteomic Approaches
Accurate quantification of hybrid E/M states requires multi-dimensional assessment of epithelial and mesenchymal characteristics at both RNA and protein levels. While transcriptomic analyses have dominated EMT research, recent evidence highlights significant discrepancies between mRNA and protein abundances due to post-translational regulation, making proteomic approaches essential for accurate phenotyping [67].
Table 2: Quantitative Methods for Assessing Hybrid E/M States
| Method Type | Specific Approach | Key Targets | Advantages | Limitations |
|---|---|---|---|---|
| Transcriptomics | RNA sequencing, RT-qPCR | EMT-TFs (SNAI1, ZEB1), E/M markers | Comprehensive, high-throughput | Poor correlation with protein levels |
| Targeted Proteomics | Parallel Reaction Monitoring (PRM) | 31+ EMT-associated proteins | Highly sensitive and quantitative | Requires specialized equipment |
| Immunofluorescence | Multiplexed imaging | E-cadherin, N-cadherin, Vimentin | Spatial context, single-cell resolution | Semi-quantitative, limited multiplexing |
| Flow Cytometry | Surface marker analysis | CD104, CD44, CD51, CD61 | High-throughput, single-cell | Limited to surface markers |
Advanced proteomic techniques, particularly parallel reaction monitoring (PRM), have enabled precise quantification of EMT-associated proteins across cancer cell lines [67]. This targeted mass spectrometry approach allows absolute quantification of proteins down to the attomole range, providing a robust method for classifying cellular phenotypes along the E/M spectrum. Studies implementing PRM have demonstrated that proteomic analysis can effectively distinguish epithelial, hybrid E/M, and mesenchymal phenotypes across diverse cancer types, with improved accuracy compared to transcriptomic approaches alone [67].
Computational Scoring and Mathematical Modeling
Mathematical models have been instrumental in understanding the dynamics and stability of hybrid E/M states. Quantitative modeling of the core EMT regulatory network has revealed that hybrid E/M states can represent stable attractors rather than transient intermediates, with cells capable of maintaining these states indefinitely under certain conditions [64]. These models capture the emergent dynamics of regulatory networks, including transcription factors, microRNAs, and phenotypic stability factors, providing a framework for predicting cellular behavior in response to genetic perturbations or microenvironmental signals [64].
Computational EMT scoring methods have been developed to position cells along the E/M spectrum based on transcriptomic or proteomic data. These approaches typically calculate scores ranging from -1 (fully epithelial) to +1 (fully mesenchymal), with hybrid E/M states occupying intermediate values [67]. Single-cell analyses have further refined these approaches, revealing continuous distributions of E/M states rather than discrete categories, highlighting the dynamic nature of epithelial-mesenchymal plasticity [67] [64].
Experimental Models and Methodologies
In Vitro Models for Hybrid EMT Analysis
The study of hybrid E/M states employs diverse experimental models that recapitulate specific aspects of epithelial plasticity in controlled settings.
Diagram: Experimental Workflow for Hybrid EMT Analysis. This workflow outlines key methodologies for inducing and characterizing hybrid E/M states in vitro, incorporating 2D, 3D, and microfluidic approaches with subsequent molecular and functional analysis.
Two-dimensional monolayer cultures represent the most accessible system for studying EMT. Treatment of epithelial cell lines (such as MDCK, MCF-10A, or A549) with EMT-inducing signals like TGF-β enables controlled induction of hybrid E/M states [34]. In the MDCK model, different sublines exhibit distinct EMT behaviors: MDCK-II cells maintain residual cell-cell adhesion with weak mesenchymal activation (stable partial EMT), while MDCK-NBL2 cells progress through partial EMT to a full mesenchymal phenotype, providing a comparative system for studying hybrid state stabilization [34]. These models enable detailed molecular characterization through transcriptomics, proteomics, and immunofluorescence, revealing the dynamic regulation of EMT-TFs and markers during transition states.
Three-dimensional culture models, including spheroids and organoids, offer more physiologically relevant contexts for studying hybrid E/M states. When embedded in collagen matrices, epithelial cells form polarized hollow spheres that, upon EMT induction, exhibit invasive protrusions while maintaining core epithelial characteristics [34]. These models particularly excel at visualizing collective cell migration, where leader cells with hybrid E/M phenotypes (elevated mesenchymal characteristics) guide follower cells with more epithelial features, recapitulating invasion patterns observed in human tumors [63]. The application of 3D models has demonstrated that hybrid E/M states are preferentially localized at the invading edge of tumors, where they coordinate multicellular invasion [63].
Methodologies for Functional Characterization
Functional characterization of hybrid E/M states requires integrated approaches that assess both molecular markers and cellular behaviors:
Wound healing assays (scratch assays) measure collective migration capabilities, where hybrid E/M cells typically exhibit intermediate migration rates compared to epithelial (low) and mesenchymal (high) phenotypes [68]. The preservation of limited cell-cell contacts in hybrid E/M states enables coordinated movement of cell sheets, distinct from the individual migration of fully mesenchymal cells.
Transwell migration and Matrigel invasion assays quantify the ability of cells to traverse membrane barriers, with or without extracellular matrix coatings [68]. Hybrid E/M cells often demonstrate enhanced invasive capabilities compared to fully mesenchymal cells, potentially due to their ability to engage both cell-matrix and residual cell-cell adhesion systems.
Matrix degradation assays evaluate proteolytic activity, a key feature of invasive hybrid E/M cells. These assays frequently detect elevated expression and activity of protease systems like urokinase plasminogen activator (uPA) and its receptor (uPAR), which activate metalloproteinases (MMPs) that degrade basement membrane components during invasion [68].
Molecular Regulation of Hybrid E/M States
Transcription Factor Networks and Signaling Pathways
The stabilization of hybrid E/M states is governed by complex regulatory networks centered on EMT transcription factors. Different EMT-TFs play distinct roles in regulating transition along the E/M spectrum. SNAIL1 acts as a pioneer factor that initiates both embryonic-like and adult-like EMT trajectories, with rapid activation following EMT induction [34]. PRRX1 specifically drives progression along the embryonic-like invasive trajectory, promoting transition to a full mesenchymal phenotype when highly expressed [34]. The dynamics of EMT-TF activation create a sequential recruitment pattern that determines positioning along the E/M spectrum, with different TF combinations stabilizing different intermediate states.
Diagram: Regulatory Network Governing Hybrid E/M States. This network illustrates the sequential activation of EMT transcription factors and their distinct roles in hybrid state stabilization versus complete EMT progression.
Multiple signaling pathways converge to regulate the core EMT network. TGF-β signaling serves as a potent inducer of EMT, activating SMAD-dependent and independent pathways that trigger EMT-TF expression [4]. Wnt/β-catenin signaling stabilizes nuclear β-catenin, which collaborates with EMT-TFs to repress epithelial genes. Inflammatory signaling through NF-κB creates a positive feedback loop with EMT-TFs, particularly ZEB1, reinforcing hybrid E/M states and creating immunosuppressive microenvironments [66]. These signaling pathways integrate inputs from the tumor microenvironment, allowing dynamic adjustment of cellular phenotype in response to contextual cues.
Epigenetic Regulation and Non-Coding RNAs
Epigenetic mechanisms play crucial roles in stabilizing hybrid E/M states by creating flexible gene expression patterns that can be maintained through cell divisions but remain responsive to microenvironmental signals. DNA methylation changes at epithelial and mesenchymal gene promoters create permissive or restrictive chromatin states that influence positioning along the E/M spectrum [4]. Histone modifications, including H3K4 methylation, H3K9 methylation, and H3K27 acetylation, dynamically regulate accessibility of EMT-related genes, with different modifications associated with epithelial versus mesenchymal chromatin states [4].
Non-coding RNAs, particularly microRNAs, serve as critical regulators of hybrid E/M plasticity. The miR-200 family and miR-34 function as EMT brakes by targeting ZEB1 and ZEB2 mRNAs, creating double-negative feedback loops that stabilize hybrid E/M states [64]. Long non-coding RNAs including HOTAIR and ZEB2NAT modulate the activity or expression of EMT-TFs, fine-tuning the positioning of cells along the E/M spectrum [63]. These regulatory RNAs respond to microenvironmental signals and contribute to the dynamic plasticity of hybrid E/M states.
Research Reagent Solutions for Hybrid EMT Studies
Table 3: Essential Research Reagents for Hybrid EMT Investigation
| Reagent Category | Specific Examples | Application | Key Functions |
|---|---|---|---|
| EMT Inducers | Recombinant TGF-β, HGF, FGF, TNF-α | Induce EMT in model systems | Activate signaling pathways that initiate EMT program |
| EMT Inhibitors | SB431542 (TGF-βR inhibitor), LY2157299 | Prevent or reverse EMT | Block upstream signaling cascades |
| Antibodies for Detection | Anti-E-cadherin, Anti-N-cadherin, Anti-Vimentin | Immunofluorescence, Western blot | Identify epithelial and mesenchymal markers |
| qPCR Assays | TaqMan assays for CDH1, CDH2, VIM, SNAI1, ZEB1 | Gene expression analysis | Quantify transcript levels of E/M markers |
| Cell Lines | MDCK, MCF-10A, NMuMG, A549 | In vitro modeling | Provide controlled systems for EMT manipulation |
| Proteomic Tools | SIL peptides, PRM assay panels | Targeted protein quantification | Absolute quantification of EMT-associated proteins |
Clinical Implications and Therapeutic Opportunities
Role in Cancer Progression and Treatment Resistance
Hybrid E/M states contribute significantly to cancer aggressiveness through multiple mechanisms. Cells in hybrid states demonstrate enhanced tumor-initiating capacity and stemness properties, making them particularly effective at founding new tumor lesions [63]. This association between hybrid E/M phenotypes and cancer stem cells (CSCs) provides a mechanistic link between cellular plasticity and tumor recurrence. Additionally, hybrid E/M cells exhibit increased therapy resistance, surviving conventional treatments that eliminate more differentiated tumor cells [63] [69]. This resistance stems from multiple factors, including enhanced DNA repair capacity, detoxification systems, and survival signaling pathways upregulated in hybrid states.
In the metastatic cascade, hybrid E/M states enable multiple aspects of dissemination. Circulating tumor cells (CTCs) frequently display hybrid E/M characteristics, co-expressing epithelial and mesenchymal markers that facilitate survival in circulation and extravasation at distant sites [63] [67]. The preservation of limited epithelial features enables cluster formation, creating cooperative units that enhance metastatic efficiency compared to individually migrating mesenchymal cells. At secondary sites, the plasticity of hybrid E/M states enables adaptation to foreign microenvironments and reversion toward epithelial phenotypes (MET) that supports metastatic outgrowth [69].
Therapeutic Targeting Strategies
Targeting hybrid E/M states presents unique challenges and opportunities for therapeutic development. Several strategic approaches show promise:
Phenotypic stability factor targeting aims to lock cells in either epithelial or mesenchymal states, reducing plasticity that drives adaptation and resistance [63] [64]. By reinforcing epithelial programs through GRHL2 activation or disrupting mesenchymal stabilization by targeting PRRX1 or ZEB1, this approach seeks to limit dynamic transitions along the E/M spectrum.
Inflammatory pathway modulation addresses the crosstalk between hybrid E/M states and the tumor immune microenvironment [66]. Hybrid E/M cells create immunosuppressive niches through cytokine and chemokine secretion, enabling immune evasion. Breaking this alliance through inflammatory pathway inhibitors may restore immune surveillance and eliminate plastic cell populations.
Metabolic targeting exploits the unique metabolic features of hybrid E/M cells, which demonstrate enhanced flexibility in fuel utilization compared to terminal states [4]. This includes dependencies on specific glycolytic enzymes, glutamine metabolism pathways, and lipid handling systems that represent vulnerabilities in plastic cell populations.
The dynamic nature of hybrid E/M states and their similarity to physiological plasticity programs present challenges for therapeutic targeting, including potential toxicity against normal stem cell populations and wound healing processes. However, the critical role of these states in driving metastasis and therapy resistance makes them compelling targets for next-generation anticancer therapies.
Hybrid E/M states represent a critical dimension of cellular plasticity that extends beyond the traditional binary conception of EMT. These dynamic intermediate states enable coordinated collective invasion, enhance stemness properties, facilitate adaptation to therapeutic challenges, and drive metastatic progression. The investigation of hybrid E/M states requires integrated approaches combining advanced molecular profiling, functional assays in physiologically relevant models, and computational modeling to decipher the regulatory networks that stabilize these plastic phenotypes.
Looking forward, key challenges include developing more precise tools to manipulate hybrid E/M states without triggering complete EMT or MET, understanding the contextual cues that determine positioning along the E/M spectrum in different tumor types, and translating this knowledge into therapeutic strategies that target the plasticity driving cancer progression. As our technical capabilities for single-cell analysis and spatial profiling advance, we will undoubtedly uncover further complexity in the E/M spectrum, revealing new opportunities for intervention against the cellular plasticity that underlies cancer mortality.
The classical view of epithelial-mesenchymal transition (EMT) as a simple binary switch between epithelial and mesenchymal states has been fundamentally challenged by recent research. It is now recognized that cells can reside in multiple intermediate or partial states along the EMT spectrum, exhibiting hybrid epithelial/mesenchymal (E/M) characteristics with distinct functional capabilities [68] [70]. This paradigm shift is particularly relevant in primitive streak research, where gastrulation involves precisely regulated EMT events that generate mesendodermal progenitors without necessarily committing to a fully mesenchymal phenotype [71].
Understanding the spectrum of EMT completion is crucial for developmental biology, cancer metastasis, and fibrosis research. In primitive streak formation, cells undergo EMT marked by cadherin switching while maintaining differentiation plasticity, demonstrating that EMT progression can be decoupled from cell fate commitment [71]. This review synthesizes current quantitative data, experimental methodologies, and conceptual frameworks for investigating partial EMT states, providing researchers with tools to dissect this complex biological process.
Quantitative Dimensions of the EMT Spectrum
Cadherin Switching Dynamics in Mesendodermal Differentiation
Recent single-cell analyses of human pluripotent stem cell (hPSC) differentiation to primitive streak lineages have revealed unexpected dynamics in cadherin expression during EMT. The degree of E-cadherin downregulation and N-cadherin upregulation varies significantly across different mesendodermal subtypes, demonstrating independent regulation of these classic EMT markers [71].
Table 1: Cadherin Expression Patterns During hPSC Differentiation to Mesendodermal Subtypes
| Cell Type | Protocol Signaling | E-Cadherin Downregulation | N-Cadherin Upregulation | Hybrid E/M State Percentage |
|---|---|---|---|---|
| Definitive Endoderm (DE) | High Activin, Low Wnt | Early (12 hours) | Moderate (24 hours) | 10-15% E-Cad+/N-Cad+ |
| Paraxial Mesoderm (PM) | Wnt activation, BMP inhibition | Early (12 hours) | High (24 hours) | 10-20% E-Cad+/N-Cad+ |
| Lateral Mesoderm (LM) | BMP activation, Wnt inhibition | Early (12 hours) | High (24 hours) | 15-25% E-Cad+/N-Cad+ |
The data reveals that E-cadherin downregulation occurs consistently at 12 hours across all primitive streak subtypes, while N-cadherin upregulation follows different timelines and magnitudes. Notably, a significant subpopulation of cells (10-25%) maintains simultaneous expression of both cadherins, representing a stable hybrid E/M state [71].
EMT Marker Correlation Analysis
The correlation between classic EMT markers varies significantly across different EMT-inducing contexts, challenging the concept of a coordinated EMT program.
Table 2: Correlation Coefficients Between EMT Markers Across Experimental Systems
| EMT Context | E-cad vs Vimentin | E-cad vs N-cad | Snail vs E-cad | ZO-1 vs Vimentin |
|---|---|---|---|---|
| Primitive Streak (hPSC) | -0.38 | -0.42 | -0.15 | -0.51 |
| Cancer Cell Lines | -0.62 | -0.71 | -0.45 | -0.68 |
| Renal Fibrosis | -0.55 | -0.58 | -0.52 | -0.61 |
| TGF-β-Induced | -0.75 | -0.82 | -0.68 | -0.79 |
The weaker correlations observed in primitive streak systems compared to cancer or fibrosis models suggest developmental EMT may involve more nuanced regulation with greater potential for intermediate states [71].
Experimental Models for Partial EMT Analysis
hPSC-Based Primitive Streak Models
The differentiation of human pluripotent stem cells (hPSCs) to primitive streak and mesendodermal subtypes provides a powerful platform for investigating EMT dynamics during development [71].
Protocol: Primitive Streak Induction and EMT Monitoring
hPSC Maintenance: Culture hPSCs in mTeSR or equivalent pluripotency maintenance medium on Matrigel-coated plates.
Primitive Streak Induction:
- Anterior PS: Activin (100 ng/mL), FGF (20 ng/mL), CHIR99021 (3 μM)
- Mid PS: Activin (50 ng/mL), BMP4 (10 ng/mL), FGF (20 ng/mL), CHIR99021 (3 μM)
- Posterior PS: BMP4 (50 ng/mL), FGF (20 ng/mL), CHIR99021 (3 μM)
Mesendodermal Specification:
- Definitive Endoderm: 48h with high Activin (100 ng/mL), BMP inhibitor (100 nM)
- Paraxial Mesoderm: 48h with CHIR99021 (3 μM), BMP inhibitor (100 nM)
- Lateral Mesoderm: 48h with BMP4 (50 ng/mL), Wnt inhibitor (1 μM)
EMT Progression Analysis:
- Timepoints: 0h, 12h, 24h, 48h
- Metrics: Flow cytometry for E-cadherin, N-cadherin, intracellular markers
- Imaging: Immunofluorescence for ZO-1, VIMENTIN, cytoskeletal organization
This system enables precise tracking of EMT dynamics while correlating with cell fate decisions, revealing that anterior primitive streak cells maintain both endoderm and mesoderm potential despite undergoing partial EMT [71].
Functional Assays for Hybrid E/M States
Several functional assays have been adapted to characterize the unique properties of cells in partial EMT states:
Invasion and Migration Assessment:
- Modified Boyden chamber assays with ECM-coated membranes
- Spheroid invasion in 3D Matrigel with quantification of invasion pattern
- Wound healing assays with single-cell tracking of migration mode
Collective vs Individual Migration Analysis:
- Time-lapse microscopy to quantify leader-follower cell behaviors
- Cell tracking software to analyze migration persistence and velocity
- Cell-cell adhesion measurements via atomic force microscopy
Cells in hybrid E/M states frequently exhibit collective migration patterns with maintained, though weakened, cell-cell junctions, distinct from either fully epithelial or fully mesenchymal cells [68] [70].
Molecular Regulation of EMT Gradients
Signaling Pathways Governing EMT Transitions
Multiple signaling pathways interact to position cells along the EMT spectrum, with different pathways predominating in various biological contexts.
Diagram 1: EMT signaling network regulating cadherin switching.
The diagram illustrates how multiple signaling pathways converge on core EMT transcription factors that coordinately regulate epithelial and mesenchymal gene expression. The balance and combination of these activated pathways determine the position along the EMT spectrum [68] [72] [73].
Metabolic and Epigenetic Regulation
Recent research has revealed additional layers of regulation controlling EMT progression:
Metabolic Reprogramming:
- Hybrid E/M states show distinct metabolic profiles with enhanced glycolysis and mitochondrial reprogramming
- Ferroptosis pathway activation can promote EMT in renal fibrosis models [74]
- Metabolic enzymes can directly regulate EMT transcription factor activity
Epigenetic Control:
- Histone modifications at epithelial and mesenchymal gene promoters create permissive or restrictive chromatin states
- DNA methylation patterns stabilize hybrid E/M states
- Non-coding RNAs fine-tune EMT progression
Technical Framework for EMT Spectrum Analysis
Research Reagent Solutions
Table 3: Essential Reagents for EMT Spectrum Research
| Reagent Category | Specific Examples | Function in EMT Research |
|---|---|---|
| Signaling Modulators | TGF-β1, BMP4, CHIR99021 (Wnt activator), Repsox (TGF-β inhibitor) | Induce or inhibit EMT progression to generate defined states along the spectrum |
| Cell Lines | hPSCs (H9, RUES2), HK-2 renal tubular cells, EMT lineage tracer lines | Provide models for developmental, fibrosis, and cancer EMT contexts |
| Antibodies | E-cadherin, N-cadherin, Vimentin, ZO-1, Snail, Twist | Quantify protein expression changes during EMT progression |
| Reporters | E-cadherin-GFP, N-cadherin-RFP, ZEB1 promoter-luciferase | Live tracking of EMT dynamics in real time |
| ECM Substrates | Matrigel, Collagen I, Laminin-511, Synthetic hydrogels | Provide microenvironmental context for EMT regulation |
Computational and Imaging Approaches
Single-Cell RNA Sequencing Analysis:
- Dimensionality reduction (UMAP, t-SNE) to visualize EMT continuum
- Pseudotime analysis to reconstruct EMT trajectories
- Cluster analysis to identify distinct E/M states
High-Content Imaging:
- Multiplexed immunofluorescence for 8+ markers
- Machine learning-based classification of cellular states
- Morphometric analysis of cell shape and cytoskeletal organization
Lineage Tracing:
- CRISPR-Cas9-based lineage recorders
- Fluorescent timer proteins
- Barcoded viral libraries to track EMT progression
Diagram 2: Experimental workflow for EMT spectrum analysis.
Implications for Primitive Streak Research and Beyond
The spectrum concept of EMT completion has profound implications for understanding primitive streak formation and differentiation. During gastrulation, cells undergo EMT to ingress through the primitive streak, but must retain precisely regulated plasticity to adopt appropriate mesendodermal fates [71].
Recent work demonstrates that anterior primitive streak cells maintain both endoderm and mesoderm potential despite undergoing EMT, while posterior primitive streak cells show restricted potential, indicating that EMT progression and fate commitment can be decoupled [71]. This challenges the traditional view that complete EMT is required for lineage specification.
The identification of stable hybrid E/M states in development suggests similar states may be relevant in other contexts:
- Cancer: Hybrid E/M states may enhance collective invasion and metastatic competence
- Fibrosis: Partial EMT in tubular epithelial cells drives collagen production while maintaining epithelial characteristics [74]
- Wound Healing: Hybrid states may optimize migration while maintaining tissue integrity
Future Directions and Technical Challenges
Several key challenges remain in investigating the EMT spectrum:
Standardization of State Definitions:
- Consensus markers and thresholds for defining hybrid E/M states
- Integration of transcriptomic, proteomic, and functional data
- Context-specific definitions for development, cancer, and fibrosis
Technical Innovation Needs:
- Live reporters for multiple EMT parameters simultaneously
- Microenvironment control to dissect ECM and mechanical influences
- High-throughput screening approaches for hybrid E/M states
Therapeutic Translation:
- Targeting specific EMT states rather than complete EMT inhibition
- State-specific vulnerabilities in hybrid E/M cells
- Biomarkers for EMT state detection in clinical samples
Understanding the spectrum of EMT completion will require continued development of sophisticated experimental models, analytical frameworks, and conceptual models that embrace the complexity of this fundamental biological process.
The epithelial-mesenchymal transition (EMT) represents a fundamental cellular process driving critical events in embryonic development, tissue repair, and cancer progression. Within the primitive streak during embryogenesis, EMT facilitates the conversion of epithelial cells into motile mesenchymal cells, enabling the complex morphogenetic events that establish the basic body plan [75] [76]. This developmental program is co-opted in pathological contexts, particularly in carcinoma progression, where it drives invasion and metastasis. The interpretation of core biomarkers like E-cadherin and Vimentin sits at the heart of understanding EMT dynamics. However, this interpretation is fraught with challenges, including cellular heterogeneity, context-dependent expression, and the spectrum of intermediate states that cells occupy during this transition. This technical guide examines these complexities within the framework of primitive streak research and its implications for modern cancer biology.
The classical view of EMT as a simple, binary switch has been substantially refined. Current consensus recognizes EMT as a dynamic, reversible process where cells can attain various stable intermediate states along the epithelial-mesenchymal spectrum [9]. This plasticity is epitomized in the primitive streak of the developing embryo, where a precise spatiotemporal control over EMT and its reverse process, MET (mesenchymal-epithelial transition), orchestrates germ layer formation and organogenesis [75]. The biomarkers used to define this process must therefore be interpreted not as simple on/off switches, but as molecular readouts of a continuous biological spectrum.
Core Biomarkers in EMT: Quantitative Expression and Functional Significance
Established Markers and Their Dynamic Regulation
The core machinery of EMT involves coordinated downregulation of epithelial markers and upregulation of mesenchymal components. The following table summarizes the key biomarkers, their functions, and expression dynamics during EMT.
Table 1: Core EMT Biomarkers and Their Characteristics
| Biomarker | Type | Primary Function | Expression in EMT | Technical Detection Methods |
|---|---|---|---|---|
| E-cadherin | Epithelial Marker | Calcium-dependent cell-cell adhesion; maintains epithelial integrity [77] | Downregulated [78] [75] [77] | IHC, ICC/IF, Western Blot [78] [77] |
| Vimentin | Mesenchymal Marker | Type III intermediate filament; maintains cell shape, cytoskeletal integrity [78] | Upregulated [78] [73] [75] | IHC, ICC/IF, Western Blot [78] [77] |
| N-cadherin | Mesenchymal Marker | Calcium-dependent adhesion; supports motility and migration [77] | Upregulated (a phenomenon known as "cadherin switch") [75] [77] | IHC, IP, Western Blot [77] |
| Transcription Factors (Snail, Slug, Twist, ZEB1) | Regulators | Transcriptional repressors of E-cadherin; inducers of mesenchymal program [75] | Upregulated [75] [9] | IHC, Western Blot, RNA-seq [75] [34] |
Quantitative Relationships in Pathological Contexts
The inverse relationship between E-cadherin and Vimentin expression is a hallmark of EMT, but this dynamic must be understood quantitatively rather than absolutely. Research in colorectal cancer (CRC) provides illustrative data:
Table 2: Correlation of E-cadherin and Vimentin with Clinicopathological Parameters in Colorectal Cancer (n=39) [78]
| Clinical Parameter | E-cadherin Expression | Vimentin Expression | Statistical Significance (p-value) |
|---|---|---|---|
| Increasing Tumor Grade | Decreased | Increased | < 0.05 |
| Advanced Pathological Stage | Decreased | Increased | < 0.05 |
| Lymph Node Metastasis | Decreased | Increased | < 0.05 |
| Vascular Invasion | Decreased | Increased | < 0.05 |
| Patient Age/Sex | No significant variation | No significant variation | > 0.05 |
This quantitative data demonstrates that biomarker interpretation must be contextualized within specific disease parameters. The expression levels correlate strongly with progression metrics but not with patient demographics, highlighting their role as disease-state indicators rather than general patient characteristics.
Key Challenges in Biomarker Interpretation
Cellular Plasticity and Heterogeneity
A primary challenge is the inherent plasticity of EMT. Cells undergoing EMT frequently reside in hybrid or partial states co-expressing both epithelial and mesenchymal markers, rather than completing a full transition [34] [9]. This heterogeneity is not biological noise but a functional characteristic of the process. In breast cancer models, researchers have identified segregated tumor cell populations activating distinct EMT trajectories—one driving invasion and another controlling inflammation [34]. Such findings necessitate a move beyond bulk analysis toward single-cell resolution to deconvolute these complex cellular landscapes.
Context-Dependent Expression and Function
Biomarker expression and functional significance vary dramatically across biological contexts. Vimentin exemplifies this complexity. While intracellular vimentin provides structural support during cytoskeletal reorganization, extracellular vimentin can function as a pro-angiogenic factor mimicking VEGF or modulate immune responses by interacting with pattern recognition receptors [73]. This multifunctionality means that detecting vimentin expression alone is insufficient; understanding its spatial distribution (intracellular vs. extracellular) and modification state is critical for accurate biological interpretation.
Technical and Methodological Variability
Discrepancies in biomarker interpretation often stem from methodological differences. Immunohistochemistry, the most common detection method, suffers from variability in antibody specificity, antigen retrieval methods, and scoring systems [78]. The field lacks universal standards for defining "loss" of E-cadherin or "gain" of vimentin, making cross-study comparisons challenging. Furthermore, fixed-tissue analyses provide static snapshots of a dynamic process, potentially missing critical transitional states.
Advanced Methodologies for EMT Analysis
Single-Cell Resolution and Trajectory Inference
Next-generation approaches leverage single-cell RNA sequencing (scRNA-seq) to map EMT trajectories with unprecedented resolution. These methods can identify transitional states and detect novel biomarkers that bulk analyses would obscure.
Table 3: Experimental Workflow for Single-Cell EMT Trajectory Analysis [79]
| Step | Method/Platform | Key Parameters | Output |
|---|---|---|---|
| 1. Cell Preparation & Sequencing | 10x Genomics, Smart-seq2 | Cell viability >80%, mitochondrial content <3% [79] | Single-cell transcriptomes |
| 2. Quality Control & Normalization | Seurat v4.1 [79] | Filter: RNA counts 100-8,000 features [79] | Normalized expression matrix |
| 3. Clustering & Dimensionality Reduction | Seurat FindNeighbors/FindClusters | Resolution parameter: 0.4-1.2 | Cell clusters (t-SNE/UMAP) |
| 4. Trajectory Inference | Slingshot v2.2 [79] | Start/end cluster definition | Pseudotemporal ordering |
| 5. Differential Expression Analysis | tradeSeq v1.8.0 [79] | Wald test (startVsEndTest) | Lineage-specific biomarkers |
This workflow has identified novel EMT-related biomarkers like PLPP4 and LTB that traditional differential expression methods overlooked [79], demonstrating the power of trajectory-based analysis for discovering transitional state markers.
Functional Invasion and Migration Assays
Complementing molecular analyses, functional assays bridge the gap between biomarker expression and phenotypic manifestation. The classical approach includes:
Transwell Invasion Assay Protocol:
- Coat Transwell inserts (8μm pore size) with Matrigel (1-2mg/mL) and allow to solidify at 37°C for 4-6 hours.
- Serum-starve cells for 24 hours, then seed 5×10⁴ cells in serum-free medium into the upper chamber.
- Place complete medium with 10% FBS as chemoattractant in the lower chamber.
- Incubate for 24-48 hours at 37°C, 5% CO₂.
- Remove non-invading cells from the upper membrane surface with a cotton swab.
- Fix invaded cells on the lower membrane surface with 4% paraformaldehyde and stain with 0.1% crystal violet.
- Count cells in 5-10 random fields under 100-200× magnification.
This assay, when correlated with E-cadherin loss and vimentin gain, provides functional validation of EMT progression [70]. Advanced 3D culture systems, organoids, and microfluidic devices now offer more physiologically relevant models for studying EMT in complex microenvironments.
Molecular Regulation and Signaling Pathways
The core EMT program is orchestrated by a network of transcription factors and signaling pathways. The following diagram illustrates the key regulatory network:
Diagram 1: Molecular Regulation of EMT (Title: Core EMT Transcriptional Network)
This regulatory network shows that SNAIL1 acts as a pioneer factor initiating both embryonic-like (invasive) and adult-like (inflammatory) EMT trajectories [34]. PRRX1 specifically drives the invasive trajectory and is required for complete mesenchymal transition [34]. These factors collectively repress epithelial genes like E-cadherin while activating mesenchymal genes like vimentin and N-cadherin.
The Scientist's Toolkit: Essential Research Reagents
Table 4: Essential Research Reagents for EMT Biomarker Investigation
| Reagent/Category | Specific Examples | Research Application | Key Considerations |
|---|---|---|---|
| Validated Antibodies | Anti-E-cadherin [EP700Y] (ab40772) [77], Anti-Vimentin [EPR3776] (ab92547) [77], Anti-N-cadherin (ab18203) [77] | IHC, ICC/IF, Western Blot | Clone validation, species cross-reactivity, application-specific optimization required |
| EMT Inducers | Recombinant TGF-β, TNF-α, EGF [79] | In vitro EMT induction in cell lines | Concentration optimization (e.g., TGF-β 2-10 ng/mL), treatment duration (hours to days) [79] |
| Cell Line Models | MDCK variants [34], DU145 (prostate) [79], A549 (lung) [79] | In vitro EMT mechanism studies | Baseline epithelial status validation, inducibility confirmation required |
| siRNA/shRNA Libraries | SNAI1-targeting, PRRX1-targeting [34] | Functional validation of EMT-TFs | Efficiency validation (>70% knockdown), off-target effect controls essential |
| Single-Cell RNA-seq Platforms | 10x Genomics, Smart-seq2 [79] | High-resolution EMT trajectory mapping | Cell quality critical (viability >80%), mitochondrial content thresholding (<3%) [79] |
Future Perspectives and Concluding Remarks
The interpretation of EMT biomarkers is evolving from a qualitative assessment of a few proteins to a quantitative, multi-dimensional analysis of cellular states. Future directions will likely include:
High-Plex Spatial Profiling: Technologies like spatial transcriptomics and cyclic immunofluorescence will enable mapping of EMT biomarkers within tissue architecture, revealing how spatial context influences transition states.
Dynamic Biosensors: Engineered reporters for EMT transcription factor activity or cell junction integrity will provide real-time monitoring of transition dynamics in live cells and organisms.
Computational Integration: Machine learning approaches that integrate multiple biomarker modalities (transcriptomic, proteomic, morphological) will generate more robust EMT state classifiers.
Therapeutic Targeting: Understanding the distinct functions of EMT trajectories may enable selective targeting of the invasive program while preserving the beneficial aspects of plasticity in tissue repair.
The challenges in interpreting E-cadherin, vimentin, and other EMT biomarkers reflect the biological complexity of the process itself. By adopting more sophisticated analytical frameworks that account for cellular plasticity, contextual signals, and dynamic transitions, researchers can extract deeper insights from these fundamental markers. The primitive streak continues to provide an essential developmental reference for understanding the principles of cellular plasticity that are repurposed in disease states, reminding us that the most profound insights often come from tracing biological pathways to their embryonic origins.
Distinguishing EMT from Other Morphogenetic Movements
Within the study of developmental biology, epithelial-mesenchymal transition (EMT) represents a fundamental differentiation switch wherein polarized epithelial cells undergo a phenotypic transformation into motile mesenchymal cells [80]. This process is crucial for numerous morphogenetic events during embryogenesis, including the formation of the primitive streak during gastrulation [2]. However, EMT constitutes just one of several morphogenetic movements that shape the embryo. It is therefore critical for researchers to distinguish the unique cellular and molecular hallmarks of EMT from other mechanisms of tissue rearrangement, such as invagination, involution, or convergent extension [2] [81]. This guide provides a technical framework for making these distinctions, with a specific focus on the context of primitive streak research, to aid scientists in accurately identifying and characterizing EMT in experimental models.
Defining EMT and Other Morphogenetic Movements
Core Characteristics of EMT
EMT is a specific cellular process characterized by a defined sequence of molecular and phenotypic changes. It begins with the disruption of intercellular adhesion complexes (tight junctions, adherens junctions, desmosomes) and the loss of apico-basal polarity [2]. This is followed by a profound cytoskeletal remodeling, where cells form apical constrictions and disorganize their basal cytoskeleton, ultimately leading to the breakdown of the basement membrane and cell ingression [2]. Cells that have undergone EMT acquire a fibroblast-like, mesenchymal morphology and gain migratory and invasive properties [9]. The reverse process, mesenchymal-epithelial transition (MET), is equally vital in development, whereby mesenchymal cells revert to an epithelial phenotype [80] [2].
A key regulator often used as a definitive marker for EMT is the transcriptional repression of E-cadherin, frequently mediated by master regulatory transcription factors like Snail, Slug (Snai2), Twist, and ZEB [80] [9] [82]. The process is orchestrated by highly conserved signaling pathways, including TGF-β, Wnt, Notch, and receptor tyrosine kinases (RTKs) such as c-Met (activated by HGF) and FGFR [80].
Other Morphogenetic Movements
In contrast to EMT, other morphogenetic movements typically involve the coordinated, large-scale migration or reshaping of entire epithelial sheets without individual cells necessarily losing their epithelial identity [2] [81].
- Ingression: Describes the internalization of individual cells from a surface epithelium. EMT is considered a specific molecular mechanism that drives cellular ingression at the primitive streak [2].
- Invagination: The local folding or buckling of an epithelial sheet inwards into a cavity, such as during the formation of the neural tube or the Drosophila ventral furrow.
- Involution: The inward rolling of an entire epithelial cell layer over a basal surface, a movement prominent in amphibian gastrulation.
- Convergent Extension: A process where a tissue narrows in one direction (converges) while elongating in a perpendicular direction (extends) through directed cell rearrangement and intercalation, crucial for axis elongation in vertebrates.
Table 1: Distinguishing Key Morphogenetic Movements
| Movement Type | Cellular Basis | Key Regulators | Primary Outcome | Example in Development |
|---|---|---|---|---|
| EMT/Ingression | Individual cells lose adhesion, become motile | Snail, TGF-β, Wnt, FGF | Generation of migratory mesenchyme | Mesoderm formation at primitive streak [2] [81] |
| Invagination | Coordinated apical constriction in cell sheet | Shroom, Rho kinase | Tissue folding | Neural tube formation [82] |
| Convergent Extension | Cell intercalation and rearrangement | Planar Cell Polarity (PCP) pathway | Tissue narrowing and elongation | Axial elongation in vertebrate embryos [81] |
EMT in Primitive Streak Formation: A Model System
The formation of the primitive streak in amniote embryos (birds and mammals) serves as a paradigmatic model for studying a developmental EMT in vivo. The primitive streak is the site where the single-layered epiblast gives rise to the three primordial germ layers—ectoderm, mesoderm, and endoderm [2].
Signaling Pathways and Molecular Regulation
The initiation of EMT at the primitive streak is guided by a precise interplay of secreted signaling factors. In the posterior region of the embryo, canonical Wnt signaling renders epiblast cells competent to respond to EMT-inducing signals [2]. This is followed by the cooperative activity of TGF-β superfamily members, notably Nodal and Vg1, which are key inducers of the primitive streak and subsequent mesoderm ingression [2]. FGF signaling is subsequently critical for maintaining the EMT regulatory network and sustaining the production of mesodermal cells [2].
These signaling cascades activate a core set of EMT transcription factors (EMT-TFs). Snail is a primary target, which directly represses the transcription of E-cadherin, thereby dismantling adherens junctions—a critical early step in EMT [80] [82]. The following diagram illustrates this core signaling network.
Core Signaling for EMT in Primitive Streak
Distinguishing Murine Primitive Streak EMT
Live imaging and cell tracking in mouse embryos have revealed that the cellular mechanisms of primitive streak formation can differ significantly from other model organisms, highlighting the need for context-specific analysis. Unlike in chick embryos, the murine primitive streak forms in situ without large-scale polonaise movements or convergence and extension of a precursor population [81]. Instead, elongation occurs via the progressive initiation of EMT, beginning in the posterior epiblast and proceeding anteriorly. The first morphological sign is the localized loss of the basal lamina, which is tightly correlated with mesoderm ingression [81]. Epiblast cells then constrict their apices and ingress via somal translocation, rather than through massive tissue rearrangements [81].
Table 2: Key Features of EMT at the Murine Primitive Streak
| Feature | Description | Experimental Evidence |
|---|---|---|
| Basal Lamina Loss | First morphological step, strictly correlated with ingression. | Immunohistochemistry for Laminin; live imaging of GFP-labeled basement membrane components [81]. |
| Apical Constriction | Ingressing cells adopt a bottle-shaped morphology. | High-resolution confocal microscopy and electron microscopy of streak sections [81]. |
| Somal Translocation | Primary mode of ingression; cell body moves into subepithelial space. | Live imaging of fluorescently labeled embryos (e.g., mT/mG) [81]. |
| Lack of Large-Scale Movement | No polonaise movements or convergence-extension prior to streak formation. | Manual cell tracking and distortion diagrams from time-lapse movies of pre-streak embryos [81]. |
Quantitative Assessment and Experimental Protocols
Transcriptomic Scoring of EMT
Moving beyond qualitative markers, several computational methods have been developed to quantify the extent of EMT on a continuous spectrum from transcriptomic data. These are crucial for identifying hybrid E/M states.
- 76-Gene Signature (76GS): Calculates an EMT score based on a weighted sum of the expression of 76 genes, with weights derived from their correlation with CDH1 (E-cadherin). Higher positive scores indicate a more epithelial phenotype [83].
- Kolmogorov-Smirnov Test (KS): This method compares the cumulative distribution functions (CDFs) of epithelial and mesenchymal gene signatures. It provides a score ranging from -1 (epithelial) to +1 (mesenchymal) [83] [84].
- Multinomial Logistic Regression (MLR): This method uses a small panel of 23 genes to calculate the probability that a sample belongs to E, M, or hybrid E/M categories, assigning a score from 0 (pure E) to 2 (pure M), with 1 indicating a maximally hybrid phenotype [83]. A key advantage of the MLR method is its ability to help distinguish between a population of "pure" hybrid E/M cells and a mixed population of E and M cells [83].
Proteomic Validation
Given the potential discordance between mRNA and protein levels, targeted proteomics offers a more functional validation of EMT states. A parallel reaction monitoring (PRM)-based targeted proteomics assay has been developed for the absolute quantification of 31 EMT-associated proteins [84]. This method involves:
- Peptide Selection: Curating proteotypic peptides for PRM analysis from established EMT markers.
- Synthetic Standards: Using synthesized heavy isotope-labeled peptides as internal standards.
- Mass Spectrometry Analysis: Running time-scheduled PRM on a triple quadrupole or hybrid mass spectrometer.
- Data Analysis: Quantifying protein abundances using software like Skyline, with limits of detection in the attomole to femtomole range [84]. This approach confirms that protein-level quantification can more accurately assess cellular phenotype and reveal heterogeneity not always apparent in transcriptomic data.
Protocol: Live Imaging and Cell Tracking in Mouse Embryos
To directly observe EMT-related cell behaviors, such as during primitive streak formation, the following protocol can be employed [81]:
- Embryo Preparation: Use genetically labeled mouse embryos (e.g., mT/mG reporter line) at pre-streak to early streak stages (E6.0-E6.5).
- Culture Conditions: Maintain embryos in specialized culture systems under controlled conditions (temperature, gas atmosphere).
- Image Acquisition: Acquire time-lapse z-stacks through the entire depth of the posterior epiblast using a confocal or light-sheet microscope at 5-10 minute intervals over 3-5 hours.
- Cell Tracking and Analysis:
- Manually or automatically track the centroids of individual cells through the time series.
- Generate tracks to visualize and quantify cell displacement and movement patterns.
- Create distortion diagrams by plotting the positions of a group of cells at the start and end of the movie to assess for convergence and extension behavior.
- Data Interpretation: The absence of large-scale swirling movements (polonaise) and anterior displacement of midline cells, coupled with the observation of progressive basal lamina breakdown and ingression, is indicative of the murine-specific EMT mechanism [81].
The workflow for this analysis is summarized below.
Workflow for Live Imaging of EMT
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for EMT and Morphogenesis Research
| Reagent / Tool | Function/Application | Example Use in Distinguishing EMT |
|---|---|---|
| mT/mG Reporter Mouse | Dual-fluorescent reporter line; labels cell membranes. Membranes are tomato red (mT) ubiquitously, but upon Cre recombination, become GFP (mG). | Visualizing and tracking cell behaviors in live mouse embryos without the need for dye injection [81]. |
| PRM Targeted Proteomics Panel | Mass spectrometry-based absolute quantification of 31 EMT-associated proteins. | Validating EMT phenotypes at the protein level, overcoming transcript-protein discordance [84]. |
| SNAI1/SNAI2 Antibodies | Immunodetection of key EMT-transcription factors. | Identifying nuclear localization of Snail or Slug as a hallmark of active EMT induction [80] [9]. |
| E-Cadherin Antibodies | Immunodetection of a core epithelial adhesion protein. | Confirming loss of epithelial character via IHC or Western blot; its loss is a cardinal feature of EMT [80] [9]. |
| TGF-β / FGF Growth Factors | Recombinant proteins to induce EMT in cell culture models. | Used in vitro to stimulate EMT and study the downstream molecular cascade [80] [34]. |
| COMET R Package | Computational tool for inferring EMT trajectories from scRNA-seq data. | Estimating inter-state transition rates and identifying genes driving EMT in a specific context [85]. |
| RhoA/ROCK Inhibitors | Chemical inhibitors of the RhoA-ROCK signaling pathway. | Probing the role of cytoskeletal remodeling and mechanical forces in regulating EMT [82]. |
Distinguishing EMT from other morphogenetic movements requires a multi-faceted approach that integrates cellular phenotyping, molecular marker analysis, and dynamic live imaging. The primitive streak offers a powerful in vivo system for such studies. Key discriminators include the loss of epithelial junctions and basal lamina, the acquisition of individual cell motility driven by a core transcriptional program, and the context-specific nature of the cellular movements involved. Employing quantitative transcriptomic and proteomic scoring methods, alongside validated experimental protocols and reagents, will enable researchers to precisely identify EMT and its intermediate states, thereby advancing our understanding of its critical role in development and disease.
Epithelial-mesenchymal transition (EMT) represents a fundamental process in development, tissue repair, and disease pathogenesis. The establishment of standardized guidelines by the EMT International Association (TEMTIA) has created a critical framework for reducing misinterpretation and promoting cross-disciplinary collaboration in EMT research. This technical guide examines the implementation of TEMTIA standards within the specific context of primitive streak research, providing researchers with standardized assessment methodologies, experimental protocols, and analytical frameworks. By adopting these unified guidelines, the scientific community can enhance reproducibility, facilitate data comparison across experimental models, and accelerate therapeutic development targeting EMT-driven processes in cancer and fibrosis.
The primitive streak serves as the quintessential model for understanding epithelial-mesenchymal transition (EMT) during embryonic development. In amniote embryos, this transient structure forms through a precisely coordinated EMT process wherein epiblast cells undergo dramatic phenotypic changes, losing epithelial characteristics and acquiring mesenchymal properties [2] [10]. EMT encompasses dynamic changes in cellular organization from epithelial to mesenchymal phenotypes, leading to functional alterations in cell migration and invasion potential [86]. This developmental process exemplifies the remarkable plasticity of epithelial cells, which can transition between states through controlled molecular reprogramming.
The TEMTIA guidelines establish a unified terminology to describe EMT processes across diverse physiological and pathological contexts [86]. These consensus standards address the pressing need for conceptual clarity in a rapidly expanding field, with over 5,700 EMT-related publications indexed in 2019 alone. For primitive streak research specifically, standardized assessment criteria are essential for accurate interpretation of the complex cellular rearrangements that establish the three germ layers during gastrulation.
Core Principles of TEMTIA Guidelines
Unified Terminology and Definitions
The TEMTIA framework establishes precise definitions for key concepts in EMT research:
EMT: A process wherein epithelial cells lose their characteristics and acquire mesenchymal phenotypes through dynamic changes in cellular organization [86]. This transition involves loss of intercellular adhesion complexes, apico-basal polarity, and basement membrane attachment, coupled with cytoskeletal reorganization and acquisition of migratory capacity [2] [4].
MET: The reverse process (mesenchymal-epithelial transition) whereby mesenchymal cells reacquire epithelial properties, a mechanism essential for subsequent developmental processes such as somite formation and organogenesis [2] [4].
Hybrid/Intermediate States: Cellular phenotypes exhibiting both epithelial and mesenchymal characteristics, now recognized as fundamental to the EMT spectrum rather than transitional artifacts [4]. Multi-omic analyses have validated these intermediate states in embryonic systems and disease models [4].
Minimum Reporting Standards
TEMTIA guidelines define essential molecular and phenotypic markers that should be reported to substantiate claims of EMT occurrence:
Table 1: Essential EMT Markers for Experimental Reporting
| Category | Epithelial Markers | Mesenchymal Markers | Functional Assays |
|---|---|---|---|
| Cell Adhesion | E-cadherin (CDH1), tight junction proteins (ZO-1), desmosomes | N-cadherin (CDH2), vimentin, fibronectin | Cell dissociation, invasion assays |
| Cytoskeletal | Cortical actin organization | Stress fibers, focal adhesions | Immunofluorescence, live imaging |
| Transcription Factors | Epithelial maintenance factors | Snail1/2, Twist, ZEB1/2, Foxa2* | Immunostaining, RNA expression |
| Functional Changes | Static behavior, apical-basal polarity | Migratory capacity, front-rear polarity | Scratch assays, time-lapse tracking |
*Note: Foxa2 acts as an epithelial gatekeeper and EMT suppressor in endoderm formation [26].
EMT Assessment in Primitive Streak Models
Mouse Model Methodologies
Mouse embryos provide a principal mammalian model for studying EMT during primitive streak formation. Live, 4-dimensional imaging combined with immunohistochemistry has revealed that the murine primitive streak develops in situ through progressive initiation of EMT without large-scale cell migration or convergence-extension behaviors [10]. The methodology for these investigations involves:
Embryo Collection and Staging: Collect embryos at precisely timed intervals (E6.5-E7.5) corresponding to pre-streak to late-streak stages. Immediately process for fixation or live imaging [10].
Immunohistochemistry Protocol:
- Fix embryos in 4% PFA for 2 hours at 4°C
- Embed in optimal cutting temperature (OCT) compound
- Section at 5-10μm thickness
- Perform antigen retrieval with citrate buffer (pH 6.0)
- Block with 5% normal serum for 1 hour
- Incubate with primary antibodies (E-cadherin, N-cadherin, Snail1, Foxa2) overnight at 4°C
- Apply fluorescent-conjugated secondary antibodies for 2 hours at room temperature
- Counterstain with DAPI and mount for imaging [10] [26]
Live Imaging and 4D Analysis: Explanted embryos cultured in DMEM/F12 medium with 50% rat serum can be imaged for up to 24 hours using confocal or light-sheet microscopy to track dynamic cellular behaviors during EMT progression [10].
Key Signaling Pathways in Primitive Streak EMT
The molecular regulation of EMT during primitive streak formation involves conserved signaling pathways that coordinate cellular transitions:
Table 2: Signaling Pathways in Primitive Streak EMT
| Signaling Pathway | Key Ligands/Receptors | Transcription Factors | Functional Role in EMT |
|---|---|---|---|
| Wnt/β-catenin | Wnt3, Wnt8c, Frizzled receptors | T (Brachyury) | Renders epiblast competent for EMT; essential for primitive streak formation [2] [26] |
| TGF-β/Nodal | Nodal, Vg1 (GDF1/3) | Smad2/3, Foxa2 | Induces primitive streak formation; drives mesendoderm specification [2] [26] |
| FGF | FGF4, FGFR1 | Snail1/2, Twist1 | Maintains EMT regulatory network; sustains mesoderm production [2] |
Figure 1: Signaling pathways regulating EMT in the primitive streak. Foxa2 (green) can suppress EMT in endoderm formation [26].
Lineage-Specific EMT Variations
Recent single-cell transcriptomic analyses have revealed fundamental differences in how germ layers undergo EMT:
Endoderm Formation: Foxa2+ definitive endoderm progenitors ingress through mechanisms of epithelial cell plasticity rather than classical EMT, maintaining E-cadherin expression while synchronously upregulating N-cadherin [26]. Foxa2 acts as an epithelial gatekeeper and EMT suppressor, shielding endoderm from undergoing complete mesenchymal transition.
Mesoderm Formation: T+ mesoderm progenitors undergo classical EMT with robust upregulation of Snail1, Twist, and other EMT transcription factors, accompanied by a complete E- to N-cadherin switch and acquisition of mesenchymal morphology [26].
Experimental Workflow for Lineage Tracing:
Figure 2: Single-cell analysis workflow for EMT lineage resolution [26].
Research Reagent Solutions
Table 3: Essential Research Reagents for Primitive Streak EMT Studies
| Reagent Category | Specific Examples | Application/Function | Considerations |
|---|---|---|---|
| Reporter Systems | Foxa2-Venus fusion (FVF), Sox17-mCherry fusion (SCF), T-GFP (Brachyury) | Time-resolved lineage labeling and tracking | Enables live imaging of cell fate decisions [26] |
| Antibodies | E-cadherin, N-cadherin, Snail1, Foxa2, Brachyury (T), Phospho-Smad2/3 | Immunohistochemistry, Western blot | Validate protein localization and expression changes [10] [26] |
| Cell Culture Models | Mouse embryonic stem cells (mESCs), Embryoid bodies | In vitro differentiation to mesoderm/endoderm | Recapitulates early developmental EMT [26] |
| Inhibitors/Agonists | Wnt pathway agonists (CHIR99021), FGF receptor inhibitors, TGF-β pathway inhibitors | Pathway manipulation and functional validation | Assess necessity/sufficiency of signaling pathways [2] |
Standardized Assessment Framework
Morphological Criteria
According to TEMTIA guidelines, EMT assessment should incorporate quantitative evaluation of these morphological parameters:
Basal Lamina Integrity: Loss of laminin and collagen IV staining precedes ingression of nascent mesoderm in mouse primitive streak [10].
Cellular Architecture: Transition from columnar epithelial morphology with apical-basal polarity to elongated, front-rear polarized mesenchymal phenotype with filopodia formation [4].
Intercellular Junctions: Disruption of tight junctions, adherens junctions, and desmosomes, replaced by punctate CDH2-mediated adhesions in migratory cells [4].
Molecular Markers Validation
TEMTIA standards emphasize multimodal validation of EMT status:
Transcriptomic Analysis: Single-cell RNA sequencing using platforms such as 10X Genomics to resolve heterogeneity in EMT states [26]. Key transcriptional changes include:
- Downregulation of epithelial genes (Cdh1, Ocln, Cldn)
- Upregulation of mesenchymal genes (Cdh2, Vim, Fn1)
- EMT transcription factor expression (Snai1, Snai2, Twist1, Zeb1)
Protein-Level Validation: Western blot and quantitative immunofluorescence to confirm translation of mRNA changes and subcellular localization of key markers [26].
Functional Assays
- Migration Capacity: Time-lapse imaging to quantify cell motility following ingression [10]
- Cell Dissociation Assays: Measure resistance to mechanical dissociation as indicator of intercellular adhesion strength
- Invasion Potential: 3D collagen gel invasion assays adapted from embryonic studies [4]
Data Interpretation and Reporting Standards
Quantitative Scoring System
Researchers should implement standardized scoring for EMT progression in primitive streak studies:
Table 4: Quantitative EMT Assessment Parameters
| Parameter | Epithelial (0 points) | Intermediate (1 point) | Mesenchymal (2 points) | Measurement Method |
|---|---|---|---|---|
| E-cadherin Localization | Continuous membrane staining | Disrupted/punctate staining | Absent membrane staining | Immunofluorescence intensity |
| Cell Shape Index | 0.1-0.3 (columnar) | 0.4-0.6 (cuboidal) | 0.7-0.9 (elongated) | Length:width ratio |
| N-cadherin Expression | Undetectable | Focal expression | Widespread expression | qRT-PCR, Western blot |
| Snail1 Nuclear Localization | Absent | Focal/weak | Strong nuclear staining | Immunofluorescence |
Contextual Considerations
TEMTIA guidelines emphasize that EMT interpretation must account for biological context:
- Developmental Stage: Precisely stage embryos, as EMT competence varies throughout primitive streak progression [10]
- Spatial Organization: Distinguish between proximal (mesoderm-forming) and distal (endoderm-forming) primitive streak regions [26]
- Cell Fate Specificity: Apply different criteria for mesoderm (complete EMT) versus endoderm (partial EMT/epithelial plasticity) lineages [26]
Implementation of TEMTIA guidelines provides an essential framework for standardizing EMT assessment in primitive streak research. By adopting unified terminology, validated experimental approaches, and quantitative reporting standards, researchers can enhance reproducibility and facilitate meaningful comparisons across studies and model systems. The recognition of lineage-specific variations in EMT mechanisms, particularly the distinction between classical EMT in mesoderm formation and epithelial plasticity in endoderm formation, highlights the sophistication of this developmental process. As single-cell technologies continue to reveal new dimensions of cellular heterogeneity during gastrulation, adherence to these consensus standards will ensure that data interpretation remains robust and biologically relevant, ultimately accelerating discoveries in development, disease mechanisms, and therapeutic interventions.
EMT Across Biological Contexts: From Development to Disease
Epithelial-mesenchymal transition (EMT) represents a fundamental cellular process driving both embryonic development and cancer progression. This review provides a comprehensive comparison between Type 1 EMT, which occurs during physiological processes including gastrulation and organogenesis, and Type 3 EMT, associated with oncogenic transformation and metastasis. We examine the distinct molecular regulators, functional consequences, and phenotypic outcomes of these EMT variants, with particular emphasis on their manifestation in primitive streak formation and carcinoma progression. Advanced research methodologies and experimental models that enable precise dissection of EMT mechanisms are detailed, providing researchers with technical insights for investigating epithelial plasticity in development and disease contexts.
Epithelial-mesenchymal transition (EMT) describes a reversible biological process in which polarized epithelial cells undergo biochemical changes to assume a mesenchymal phenotype, characterized by enhanced migratory capacity, invasiveness, and resistance to apoptosis [87]. EMT is not a single uniform program but rather encompasses distinct variants with different functional consequences. The current classification system organizes EMT into three subtypes: Type 1 EMT is associated with implantation, embryogenesis, and organ development; Type 2 EMT occurs in wound healing, tissue regeneration, and organ fibrosis; and Type 3 EMT takes place in neoplastic cells that have undergone genetic and epigenetic alterations [87]. This review focuses specifically on comparing Type 1 and Type 3 EMT, as they represent fundamentally different biological programs co-opting similar molecular machinery - one for physiological morphogenesis and the other for pathological progression.
The primitive streak, a transient embryonic structure that appears during gastrulation, provides a quintessential model for studying Type 1 EMT [4] [11]. In amniotes, the primitive streak forms at the posterior region of the embryonic epiblast, where epiblast cells undergo EMT, ingress, and give rise to the mesoderm and endoderm germ layers [11]. This developmental EMT is characterized by precise spatial and temporal regulation, resulting in well-orchestrated morphogenetic movements that establish the basic body plan. In contrast, Type 3 EMT occurs in carcinoma cells that have acquired genetic mutations, enabling them to dissociate from primary tumors, invade surrounding tissues, and ultimately metastasize to distant organs [88] [87]. While both processes share common molecular features, including downregulation of epithelial markers and upregulation of mesenchymal markers, their regulatory contexts and functional outcomes differ substantially.
Comparative Analysis of Type 1 and Type 3 EMT
The following tables provide a systematic comparison of the defining characteristics, molecular regulation, and functional properties of Type 1 and Type 3 EMT.
Table 1: Fundamental characteristics of Type 1 and Type 3 EMT
| Characteristic | Type 1 EMT (Developmental) | Type 3 EMT (Oncogenic) |
|---|---|---|
| Biological context | Embryogenesis, organ development, primitive streak formation | Cancer progression, metastasis |
| Functional purpose | Tissue morphogenesis, generation of cell diversity | Local invasion, metastatic dissemination |
| Relationship with inflammation | Not associated with inflammatory processes | Often associated with inflammatory microenvironment |
| Temporal regulation | Precisely timed and transient | Dysregulated and sustained |
| Tissue outcomes | Normal tissue architecture and function | Tissue destruction, organ failure |
| Developmental potential | Can generate primary mesenchyme with MET capability | Limited differentiation potential |
| Genetic stability | Genetically stable programs | Accumulated genetic and epigenetic alterations |
Table 2: Molecular markers and regulators in Type 1 vs. Type 3 EMT
| Molecular Component | Type 1 EMT | Type 3 EMT |
|---|---|---|
| Key transcription factors | Snail1, Snail2 (Slug), Twist | Snail, ZEB, TWIST families |
| Signaling pathways | Wnt, Nodal, BMP, Fgf | TGF-β, Wnt, Notch, PI3K-AKT |
| Epithelial markers lost | E-cadherin, cytokeratins | E-cadherin, occludin, cytokeratin |
| Mesenchymal markers gained | N-cadherin, vimentin, fibronectin | N-cadherin, vimentin, fibronectin |
| ECM remodeling enzymes | Precisely regulated MMPs | Dysregulated MMPs (MMP-2, MMP-9, MMP-14) |
| Developmental regulators | Axial mesoderm determinants | Oncogenic drivers (e.g., Six1, Cripto) |
| Metabolic reprogramming | Developmental metabolic shifts | Glycolysis, metabolite plasticity |
Molecular Regulation and Signaling Networks
Transcription Factor Networks in EMT
The core transcriptional machinery of EMT is orchestrated by several families of transcription factors, including the SNAIL, ZEB, and TWIST families [88]. While these regulators participate in both Type 1 and Type 3 EMT, their expression patterns and functional hierarchies differ significantly between the two contexts.
In Type 1 EMT during gastrulation, Snail1 is activated as cells ingress at the primitive streak, where it downregulates E-cadherin transcription [11]. However, genetic studies in mouse embryos reveal that Snail1 is not absolutely required for the ingression process itself, but rather regulates subsequent mesoderm migration [11]. Similarly, in chick embryos, Snail2 (Slug) participates in gastrulation EMT, though its precise role remains to be fully elucidated [11]. The sequential activation of EMT-TFs follows a precise developmental program, with different factors recruited at specific stages of the epithelial-to-mesenchymal spectrum [34].
In Type 3 EMT, the same transcription factors are often dysregulated. SNAIL1 acts as a pioneer factor at the base of EMT trajectories, with PRRX1 driving progression toward an invasive phenotype [34]. Cancer cells can activate distinct EMT trajectories - embryonic-like programs driving invasion or adult-like programs promoting inflammation [34]. This bifurcation is not observed in developmental EMT, reflecting the dysregulated nature of oncogenic EMT programs. The ZEB family of zinc finger transcription factors (ZEB1 and ZEB2) further contributes to Type 3 EMT by binding E-box regulatory sequences to repress epithelial genes [88].
Signaling Pathway Activation
Multiple evolutionarily conserved signaling pathways initiate and modulate EMT programs in both developmental and oncogenic contexts.
TGF-β signaling represents one of the most potent inducers of EMT across biological contexts [34]. In development, TGF-β family members including Nodal and BMP direct cell fate specification and morphogenetic movements. In cancer, TGF-β signaling undergoes a paradoxical switch from tumor-suppressive to pro-oncogenic functions, directly stimulating EMT and metastasis [88].
Wnt/β-catenin signaling is essential for primitive streak formation in amniote embryos [11]. In the mouse embryo, gastrulation begins with formation of the primitive streak at the posterior region, initiated by a network of signaling pathways including Wnt, Nodal, BMP, and Fgf [11]. In cancer, dysregulated Wnt signaling promotes EMT through stabilization of β-catenin and subsequent transcriptional activation of EMT-TFs [88].
Other signaling pathways including Notch, Hedgehog, and receptor tyrosine kinase pathways contribute to EMT regulation in a context-dependent manner. In development, these pathways often act in concert to coordinate complex morphogenetic events. In cancer, these same pathways are frequently mutated or dysregulated, contributing to the aberrant activation of EMT programs.
Figure 1: Signaling networks regulating Type 1 and Type 3 EMT. While both EMT types share common regulatory nodes including core transcription factors and ECM remodeling enzymes, they are activated by distinct upstream signals and drive different biological processes.
Epigenetic and Post-translational Regulation
EMT regulation extends beyond transcriptional control through epigenetic mechanisms including DNA methylation, histone modifications, non-coding RNAs, chromatin remodeling, and 3D genome architecture [4]. These regulators orchestrate transcriptional plasticity across the EMT spectrum, enabling transitions to hybrid epithelial-mesenchymal (E/M) states or fully mesenchymal phenotypes.
In development, epigenetic regulation ensures precise spatial and temporal control of EMT programs. For example, during gastrulation, epigenetic modifications help coordinate the sequential activation of EMT transcription factors in the primitive streak [4]. In cancer, epigenetic alterations contribute to the dysregulated and sustained EMT activation characteristic of Type 3 EMT. Similarly, post-translational modifications including phosphorylation, ubiquitination, and sumoylation fine-tune the stability and activity of EMT regulators in both contexts [88].
Non-coding RNAs, particularly microRNAs, play crucial roles in EMT regulation. The miR-200 family and miR-34 function as negative regulators of EMT by targeting SNAIL and ZEB family members [88]. In development, these miRNAs help maintain epithelial states and ensure proper timing of EMT events. In cancer, downregulation of these EMT-suppressing miRNAs contributes to the activation of Type 3 EMT programs.
Functional Consequences and Phenotypic Outcomes
Cellular and Tissue-Level Changes
The functional consequences of Type 1 and Type 3 EMT differ fundamentally despite shared cellular machinery. Type 1 EMT generates cellular diversity and enables morphogenetic movements essential for proper embryonic development [87]. Cells undergoing Type 1 EMT typically retain the capacity for MET, allowing for subsequent epithelial reorganization during organogenesis. Importantly, Type 1 EMT does not cause fibrosis or induce invasive phenotypes resulting in systemic spread [87].
In contrast, Type 3 EMT enables local invasion and metastatic dissemination of carcinoma cells [87]. Cancer cells undergoing Type 3 EMT acquire stem-like properties, enhanced resistance to apoptosis, and increased therapeutic resistance [88]. Unlike the precisely regulated and self-limiting nature of Type 1 EMT, Type 3 EMT is characterized by dysregulation and persistence, contributing to disease progression and mortality.
The concept of EMT as a binary process has been revised in recent years, with both developmental and oncogenic EMT now understood as multi-step processes generating intermediate hybrid epithelial/mesenchymal states [4]. In development, these intermediate states enable precise control of morphogenetic movements. In cancer, hybrid E/M states are associated with enhanced plasticity and metastatic competence [88].
Metabolic Reprogramming
Both Type 1 and Type 3 EMT involve significant metabolic reprogramming, though with distinct regulatory constraints and outcomes. During embryonic development, metabolic adaptations support the energetic and biosynthetic demands of rapid proliferation and morphogenesis. Cancer cells similarly adapt their metabolism through different mechanisms, exhibiting metabolic plasticity that enables utilization of the same metabolite across various progression stages, or metabolic flexibility that allows exploitation of different metabolites [4].
Glycolytic reprogramming is a hallmark of both developmental and oncogenic EMT programs. However, in Type 1 EMT, metabolic changes are precisely regulated and integrated with developmental programs, while in Type 3 EMT, metabolic alterations are often dysregulated and contribute to the aggressive phenotype of carcinoma cells.
Research Methodologies and Experimental Models
The Scientist's Toolkit: Essential Research Reagents
The following table compiles key research reagents and experimental tools for investigating Type 1 and Type 3 EMT.
Table 3: Essential research reagents for EMT investigation
| Reagent Category | Specific Examples | Research Application |
|---|---|---|
| EMT Inducers | TGF-β, FGF, EGF, HGF, Wnt3a | Induction of EMT in cell culture models |
| Signaling Inhibitors | SB431542 (TGF-βR inhibitor), LY294002 (PI3K inhibitor) | Pathway-specific inhibition studies |
| Cell Line Models | MDCK, NMuMG, MCF-10A; Cancer cell lines with varying EMT states | In vitro EMT modeling and screening |
| Antibody Panels | E-cadherin, N-cadherin, vimentin, fibronectin, ZO-1 | Immunofluorescence and Western blot detection |
| qPCR Assays | Epithelial (CDH1, KRTT19) and mesenchymal (CDH2, VIM, FN1) markers | Quantitative gene expression analysis |
| 3D Culture Matrices | Matrigel, collagen I, laminin-rich ECM | Modeling tissue context and invasion |
| Transcription Factor Assays | Snail, Slug, Twist, ZEB1 reporter constructs | Monitoring TF activity and regulation |
Experimental Approaches for EMT Investigation
Multiple experimental approaches have been developed to investigate the mechanisms and functional consequences of EMT in both developmental and oncogenic contexts.
In vitro models include classical two-dimensional cell culture systems, which allow controlled manipulation of signaling pathways and straightforward assessment of molecular changes. However, these systems poorly recapitulate the tissue context of EMT. Three-dimensional culture models, including spheroids and organoids, provide more physiologically relevant environments for studying EMT [70]. These systems better mimic tissue architecture, cell-ECM interactions, and paracrine signaling networks that influence EMT programs.
Migration and invasion assays represent fundamental tools for assessing the functional consequences of EMT. Wound healing (scratch) assays measure two-dimensional cell migration, while transwell migration and Matrigel invasion assays quantify traversing capacity through porous membranes with or without ECM barriers [70]. These assays have demonstrated that molecules like u-PA/u-PAR or MMPs significantly increase migration and invasion when EMT is induced [70].
Advanced imaging approaches enable detailed characterization of EMT dynamics in developing embryos and tumor models. Live-cell imaging captures the temporal progression of EMT, while intravital imaging allows observation of EMT events within native tissue environments [11]. These approaches have revealed that gastrulation EMT in mouse embryos occurs through stochastic ingression of individual cells at the primitive streak, contrary to earlier models suggesting coordinated mass migration [11].
Lineage tracing techniques utilizing Cre-lox or similar genetic labeling systems enable definitive identification of cells that have undergone EMT in vivo. These approaches are particularly valuable for distinguishing genuine EMT events from other sources of mesenchymal cells in both developmental and tumor contexts [87].
Figure 2: Experimental workflow for EMT investigation. Research approaches span in vitro models for mechanistic studies, in vivo systems for physiological context, and advanced analytical methods for comprehensive characterization.
Analytical Methods for EMT Characterization
Comprehensive characterization of EMT requires integration of multiple analytical approaches. Molecular profiling techniques assess changes in gene expression (RNA sequencing), protein abundance and modification (proteomics), and metabolic fluxes (metabolomics) associated with EMT progression [89]. Single-cell RNA sequencing has been particularly transformative, revealing the heterogeneity of EMT states and enabling reconstruction of EMT trajectories at unprecedented resolution [34].
Computational approaches including machine learning algorithms facilitate identification of EMT signatures from complex molecular datasets [89]. For example, XGBoost models have identified MMP3, MMP9, and MT1-MMP (MMP14) transcripts and proteomic features as predictors of invasion and poor prognosis in breast cancer [89]. These computational tools enable researchers to extract meaningful biological insights from high-dimensional data generated by modern profiling technologies.
Type 1 and Type 3 EMT represent distinct biological programs that co-opt similar molecular machinery for fundamentally different purposes. Type 1 EMT drives physiological processes including gastrulation and organogenesis, is precisely regulated in space and time, and contributes to normal development. In contrast, Type 3 EMT enables pathological cancer progression, is dysregulated and sustained, and drives metastasis and therapeutic resistance. Understanding both the shared mechanisms and unique features of these EMT variants provides crucial insights into normal development and disease pathogenesis, while informing the development of novel therapeutic strategies targeting EMT in cancer. Future research efforts should focus on elucidating the context-specific regulation of EMT programs and developing interventions that can selectively target pathological EMT without disrupting its physiological functions.
Epithelial-mesenchymal transition (EMT) is a fundamental cellular process that is essential for the morphogenesis of complex tissues in metazoans. Through this evolutionarily conserved, multi-stage process, epithelial cells lose their apical-basal polarity and cell-cell adhesion, acquiring migratory capacity and mesenchymal characteristics [4] [9]. While EMT operates in various physiological and pathological contexts, two of the most significant and well-studied developmental EMT events occur at the primitive streak during gastrulation and in the neural crest during neural tube formation [65]. These primary EMTs are crucial for generating cellular diversity and establishing the basic body plan in amniotes. This review provides a detailed functional comparison of these two processes, examining their distinct roles, molecular regulation, and cellular behaviors within the broader context of embryonic development.
Developmental Context and Physiological Roles
Primitive Streak EMT in Gastrulation
Gastrulation is a critical morphogenetic process in early embryogenesis wherein a single-layered epiblast is transformed into a trilaminar structure consisting of the three primary germ layers: ectoderm, mesoderm, and endoderm [4] [65]. In amniotes (birds, reptiles, and mammals), this process is centered around the primitive streak, a linear structure that bisects the embryo along the antero-posterior axis.
- Germ Layer Formation: Cells of the epiblast move to the midline, undergo EMT at the primitive streak, and internalize to generate the mesoderm and endoderm. The cells that remain in the epiblast become the ectoderm [65].
- Evolutionary Conservation: The primitive streak and its associated EMT are evolutionarily conserved, though the specific mode of gastrulation can vary between species. Remarkably, even a single-gene knockout can modify the gastrulation mode while still achieving the same final body plan [4].
- Foundation for Organogenesis: The mesodermal and endodermal cells generated through this EMT subsequently undergo mesenchymal-epithelial transitions (MET) to form epithelia of various organ systems, including the renal, cardiovascular, and gastrointestinal systems [90].
Neural Crest EMT in Neural Tube Formation
Following gastrulation, a second major primary EMT occurs during the formation of the vertebrate nervous system. The neural crest is a transient, multipotent population of cells that originates from the dorsal aspect of the neural tube.
- Origin from Neural Ectoderm: Neural crest cells are derived from the epithelial cells of the neural plate, which rolls up to form the neural tube. Cells located near the dorsal midline undergo EMT to become migratory neural crest cells [65].
- Extensive Migration and Diversification: These cells subsequently delaminate from the neural tube and navigate along stereotypic pathways throughout the embryo, giving rise to an astonishing array of cell types and structures [65].
- Vertebrate Innovation: The neural crest is considered an evolutionary novelty that is crucial for the formation of a complex peripheral nervous system and the generation of the vertebrate head [65].
Table 1: Comparative Physiological Roles of Primitive Streak and Neural Crest EMT
| Feature | Primitive Streak EMT | Neural Crest EMT |
|---|---|---|
| Developmental Stage | Early embryogenesis (Gastrulation) | Later embryogenesis (Neurulation) |
| Embryonic Origin | Epiblast | Neural Ectoderm |
| Primary Physiological Role | Formation of mesoderm and endoderm; establishment of trilaminar embryo | Generation of migratory cells for peripheral nervous system, craniofacial structures, and pigment cells |
| Developmental Context | Ancient process, fundamental for all metazoans | Evolutionary novelty, crucial for vertebrate complexity |
| Fate of Mesenchymal Cells | Contribute to internal tissues and organs; often undergo subsequent MET | Migrate extensively; differentiate into diverse cell types at target locations |
Molecular Regulation and Signaling Pathways
The molecular orchestration of EMT involves a complex network of signaling pathways and transcription factors. While there is some overlap in the core machinery, primitive streak and neural crest EMT are distinguished by specific inductive signals and transcriptional hierarchies.
Core EMT Transcription Factors
A conserved set of EMT-transcription factors (EMT-TFs) drives the loss of epithelial characteristics and the acquisition of the mesenchymal phenotype. However, their specific expression and functional hierarchies differ between the two contexts.
- Snail Family (Snail/Snai1, Slug/Snai2): These are pivotal pioneer factors in both primitive streak and neural crest EMT. In the chick embryo, Slug was identified as a critical regulator of gastrulation [4]. Similarly, Snail and Slug are both expressed during neural crest development, with Snail acting as a pioneer factor for the process [34].
- Twist and PRRX1: These factors are often associated with more advanced or invasive EMT states. In model systems, PRRX1 is specifically required for the progression to a full, invasive mesenchymal phenotype, a characteristic feature of migratory neural crest cells [34]. Its expression pattern aligns with the invasive trajectory in the neural crest.
- Sequential Activation: Studies in neural crest development reveal a sequential activation of EMT-TFs, with Prrx1 being activated at advanced stages to stabilize the mesenchymal phenotype in migratory cells [34].
Signaling Pathways
The initiation of EMT is triggered by extracellular signals from the microenvironment. The specific combination of signals differs between the primitive streak and neural crest.
- Primitive Streak Inducers: The formation of the primitive streak and the ensuing EMT are coordinated by a combination of growth factors, including Wnts, Fibroblast Growth Factors (FGFs), and Bone Morphogenetic Proteins (BMPs) [4] [91]. Gradients of these signals, particularly BMP, help establish the proximal-posterior identity of the epiblast and prime cells for ingression through the streak [91].
- Neural Crest Inducers: Neural crest EMT is induced by signals emanating from the adjacent tissues, including BMPs, Wnts, and FGFs from the non-neural ectoderm and the paraxial mesoderm [65]. The precise spatiotemporal coordination of these signals is essential for the delamination of neural crest cells from the dorsal neural tube.
Diagram 1: Comparative signaling pathways and transcription factor hierarchies in Primitive Streak and Neural Crest EMT.
Cellular Dynamics and Functional Outcomes
The cellular behaviors and ultimate fate of the mesenchymal cells produced by primitive streak and neural crest EMT are markedly different, reflecting their distinct roles in development.
Ingression and Migration Patterns
- Primitive Streak EMT: The EMT at the primitive streak facilitates the ingression of epiblast cells into the interior of the embryo. The migration is typically a short-range movement from the streak to specific positions where the mesodermal and endodermal layers are established [4] [65]. The process involves a profound remodeling of cell adhesion, with a switch from E-cadherin to N-cadherin expression in the emerging mesenchymal cells [4].
- Neural Crest EMT: Neural crest cells undergo delamination from the dorsal neural tube and embark on extensive, long-distance migration throughout the embryo. They follow precise, conserved pathways to reach diverse target sites [65]. This process requires a robust activation of invasive programs, including the expression of proteases like matrix metalloproteinases (MMPs) and the urokinase plasminogen activator (uPA) system to degrade the extracellular matrix [68] [34].
Potency and Differentiation Potential
The mesenchymal cells generated by these two EMTs also differ in their developmental potential.
- Primitive Streak-Derived Mesenchyme: The mesoderm formed through primitive streak EMT is a highly heterogeneous population containing precursors for a vast number of cell types, including connective tissue, skeletal and muscle tissues, vascular endothelial cells, and the stromal cells of internal organs [90]. It is a fallacy, however, to refer to this early mesenchyme simply as "embryonic connective tissue," as its potential is far broader [90].
- Neural Crest-Derived Mesenchyme: The neural crest is a classic example of a multipotent stem/progenitor cell population in the embryo. Neural crest cells give rise to neurons and glia of the peripheral nervous system, melanocytes, craniofacial cartilage and bone, and connective tissue of the head [65]. This immense differentiation capacity underscores its importance in vertebrate development.
Table 2: Comparative Cellular Dynamics and Functional Outcomes
| Aspect | Primitive Streak EMT | Neural Crest EMT |
|---|---|---|
| Primary Cellular Behavior | Ingression into the embryo | Delamination and long-range migration |
| Migration Pattern | Short-range, collective movements | Extensive, individual cell migration along stereotypical paths |
| Invasion Signature | Not typically associated with a strong invasive program | Strong invasive signature (e.g., PRRX1-dependent, protease activity) [34] |
| Developmental Potency | Heterogeneous progenitors for internal organ systems | Multipotent stem/progenitor cells for diverse derivatives |
| Key Molecular Features | Cadherin switch (E- to N-cadherin) [4] | Activation of pro-invasion genes (BC-PINGs), FAK signaling [34] |
| Subsequent Transitions | Frequently followed by MET (e.g., in somite, organ formation) | Terminal differentiation at target sites, with limited MET |
Experimental Models and Methodologies
Studying the dynamics of EMT in development requires sophisticated in vitro and in vivo models that can capture the complexity and spatiotemporal regulation of the process.
In Vitro Models
- Epiblast Stem Cells (EpiSCs): Derived from the post-implantation epiblast, EpiSCs provide a valuable in vitro model for studying early lineage commitment, including the events leading to primitive streak formation [91]. These cells can be differentiated towards mesodermal and endodermal fates in response to WNT and BMP signaling.
- Neural Crest Differentiation Models: Pluripotent stem cells (ESCs/iPSCs) can be directed to differentiate into neural crest cells through defined protocols. These models allow for the molecular dissection of neural crest specification, delamination, and migration.
- 3D Organoid and Collagen Cultures: To study invasive behaviors, epithelial cells can be embedded in three-dimensional (3D) collagen matrices. For instance, MDCK-NBL2 cells treated with TGF-β in 3D collagen display protrusive activity and invasion, a phenotype dependent on PRRX1, mimicking the invasive EMT of the neural crest [34].
In Vivo and Ex Vivo Models
- Genetically Engineered Mouse Models (GEMMs): The use of orthogonal recombinase systems (e.g., Cre/Flippase) allows for precise fate-mapping of cells expressing mesenchymal markers like Vimentin, enabling the tracking of EMT-derived lineages in vivo [92]. This is powerful for analyzing both developmental and cancer-associated EMT.
- Chick Embryo Studies: The chick embryo is a classical and highly accessible model for studying both primitive streak and neural crest development due to its ease of manipulation and imaging. Electroporation of expression constructs or morpholinos enables functional gain- and loss-of-function studies [65].
- Ex Vivo Assays: Functional assays such as the chorionallantoic membrane (CAM) assay, cell line-derived xenografts, and syngeneic mouse models are instrumental in investigating intravasation, site-specific metastasis, and treatment response, processes that reactivate developmental EMT programs [68].
The Scientist's Toolkit: Key Research Reagents and Assays
Table 3: Essential Reagents and Methodologies for EMT Research
| Reagent / Assay | Function / Application | Contextual Example |
|---|---|---|
| FGF/ERK Pathway Inhibitors (e.g., PD03) | To modulate ERK signaling and study its role in epiblast priming and lineage bias [91]. | Primitive Streak Models |
| TGF-β Cytokine | A potent inducer of EMT; used to trigger transition in various epithelial cell lines [34]. | Neural Crest & Cancer Models |
| 3D Collagen Matrices | To provide a physiologically relevant environment for studying cell invasion and protrusive activity [34]. | Invasion Assays |
| FAK Inhibitors (FAKi) | To block Focal Adhesion Kinase signaling and assess its role in invasive EMT progression [34]. | Neural Crest & Cancer Models |
| Lineage Tracing Models (e.g., Vimentin-FSF-tdTomato) | To fate-map and track the progeny of cells that have undergone EMT in vivo [92]. | In Vivo Fate Mapping |
| scRNA-seq & scGET-seq | For single-cell resolution of transcriptional heterogeneity and genomic instability in EMT-derived populations [92]. | Molecular Profiling |
| Wound Healing / Scratch Assay | A classic, simple method to assess collective cell migration in two dimensions [68]. | Migration Analysis |
| Transwell Migration/Invasion Assay | To quantify the migratory and Matrigel-invasive capacity of individual cells [68]. | Metastasis Research |
The functional comparison between primitive streak EMT and neural crest EMT reveals a fascinating picture of how a conserved cellular process can be adapted to fulfill distinct developmental needs. The primitive streak EMT is primarily a generative event, focused on creating the foundational germ layers from which the majority of internal tissues and organs will form. Its EMT is a means to an end, often quickly followed by MET to build new epithelial structures. In contrast, neural crest EMT is fundamentally disseminative and transformative, dedicated to producing a migratory population of multipotent cells that will colonize distant regions of the embryo and generate a vast array of derivative cell types. This is reflected in its robust activation of invasive transcription factors like PRRX1 and its associated molecular programs.
Understanding the nuances of these developmental EMT programs is not merely an academic exercise. It provides critical insights into the mechanisms of cell plasticity that are reactivated in pathological conditions such as organ fibrosis and cancer metastasis [4] [92] [34]. The molecular signatures defining the invasive neural crest EMT, for instance, are often re-deployed by carcinoma cells to disseminate from the primary tumor. Therefore, continued research into these fundamental developmental processes, using the advanced experimental tools outlined here, will undoubtedly yield deeper insights into both embryogenesis and human disease.
Linking Gastrulation to Cancer Metastasis and Therapy Resistance
The study of gastrulation, particularly the formation of the primitive streak and the accompanying epithelial-to-mesenchymal transition (EMT), provides a foundational framework for understanding the mechanisms driving cancer metastasis and therapy resistance. The primitive streak is the site in the developing embryo where epiblast cells undergo EMT, ingressing to form the mesoderm and endoderm germ layers. This developmental program is recapitulated in carcinoma cells, which co-opt the same molecular machinery to dissociate from primary tumors, invade surrounding tissues, and disseminate to distant organs [93] [70]. Within this context, research into the primitive streak offers more than just an analogy; it reveals the core regulatory circuits that, when reactivated in cancer, drive the lethal cascade of metastasis and treatment failure. This whitepaper delineates the molecular parallels between gastrulation and metastasis, quantitative genetic evidence, emerging therapeutic strategies targeting these pathways, and detailed experimental methodologies for researchers investigating this critical intersection of developmental and cancer biology.
Biological Foundations: EMT in Development and Cancer
EMT and Cell Plasticity in Gastrulation
Contemporary research has refined the classical view that all germ layer formation involves a complete EMT followed by a mesenchymal-to-epithelial transition (MET). High-resolution single-cell transcriptomic studies of mouse embryos reveal that while the formation of the mesoderm is driven by a classical, Snail1-dependent EMT, the definitive endoderm is formed through a distinct mechanism characterized by epithelial cell plasticity [26]. In this pathway, Foxa2 acts as an EMT suppressor and epithelial gatekeeper, allowing cells to ingress without fully committing to a mesenchymal fate. These cells maintain E-cadherin expression and do not undergo a subsequent MET, as they never fully relinquish their epithelial characteristics [26] [93]. This challenges the established EMT-MET cycle paradigm for endoderm formation and underscores the spectrum of cellular plasticity available during morphogenesis.
The mechanics of ingression at the primitive streak involve a ratchet-like apical constriction process. Live imaging of mouse embryos shows that ingressing epiblast cells constrict their apices in a pulsed, stepwise fashion through the asynchronous shrinkage of apical junctions [6]. This process is regulated by an anisotropic distribution of actomyosin networks and Crumbs2 complexes at apical cell junctions. The loss of Crumbs2 disrupts myosin II localization and function, impairing ingression and highlighting its critical role as a regulator of actomyosin dynamics during this developmental EMT [6].
Reactivation of Embryonic Programs in Metastasis
The molecular programs governing gastrulation are aberrantly reactivated in carcinoma cells to execute the multi-step process of metastasis. This involves local breakdown of the basement membrane, loss of cell polarity, and induction of EMT, enabling cells to detach, invade, and intravasate into the circulation [94] [70]. Crucially, the "seed and soil" hypothesis posits that successful metastasis requires not only the disseminated tumor cell (the "seed") but also a receptive microenvironment in the distant organ (the "soil") [95]. This soil can be preconditioned by factors from the primary tumor, creating a pre-metastatic niche, a concept that mirrors the preparation of developmental fields during embryogenesis.
The role of the tumor microenvironment and the gut microbiome has emerged as a significant modulator of this process. Dysbiosis, or microbial imbalance, can promote metastasis by inducing EMT, shaping the immune landscape, and altering the bone microenvironment to foster a pre-metastatic niche [96] [97] [95]. For instance, Fusobacterium nucleatum is not only enriched in colorectal tumors but its presence is maintained in liver metastases, suggesting it actively participates in metastatic progression [95].
Quantitative Genetic Parallels
A compelling body of evidence demonstrates a significant genetic overlap between gastrulation and metastasis. A systematic analysis identified at least 50 common genes critical for both processes, wherein genetic inhibition disrupts gastrulation in model embryos, while ectopic expression promotes metastasis in cancer cells [94].
Table 1: Selection of Key Genes Involved in Both Gastrulation and Metastasis
| Gene | Role in Gastrulation | Effect in Metastasis |
|---|---|---|
| SNAI1 | Convergence and extension [94] | Induces EMT [94] |
| TWIST1 | Mesoderm formation [94] | Induces EMT [94] |
| BMP | Convergence and extension [94] | Promotes EMT [94] |
| WNT | Convergence and extension [94] | Enhances migration and invasion [94] |
| ZEB1 | Epiboly progression [94] | Induces EMT [94] |
| RHOA | Convergence and extension [94] | Promotes migration and invasion [94] |
| MMP14 | Convergence and extension [94] | Facilitates invasion [94] |
| FGFR | Convergence and extension [94] | Promotes invasion [94] |
These genes are central to signaling pathways such as TGF-β, Wnt, and FGF, and regulate core EMT transcription factors and cytoskeletal components. Their dual roles underscore gastrulation as a conserved functional template for metastatic behavior.
Implications for Therapy Resistance and Emerging Therapeutics
The cellular plasticity afforded by EMT is a key driver of therapy resistance. Cells that have undergone EMT often exhibit enhanced stemness and survival capabilities, making them refractory to conventional chemotherapy and targeted therapies [70] [98]. Consequently, targeting the EMT process itself represents a promising frontier for oncology.
Table 2: Emerging Therapeutic Strategies Targeting EMT-Related Pathways
| Therapeutic Target/Approach | Mechanism of Action | Development Status |
|---|---|---|
| Netrin-1 Inhibition | Therapeutic antibody blocks Netrin-1/UNC5B interaction, inhibiting EMT and sensitizing tumors to chemotherapy [98]. | Proof-of-concept in preclinical models and early clinical trials for endometrial cancer [98]. |
| Gut Microbiota Modulation | Probiotics, prebiotics, and FMT restore microbial balance, potentially reducing EMT-inducing signals and inflammation via the gut-immune-bone axis [96] [97]. | Preclinical and ongoing clinical trials for bone metastasis prevention [96]. |
| Phenotypic Screening | Uses zebrafish gastrulation (epiboly) as a surrogate to identify anti-metastasis drugs that interrupt conserved morphogenetic movements [94]. | Identified adrenosterone and pizotifen as hits; validated in mouse metastasis models [94]. |
The development of an anti-Netrin-1 antibody is particularly notable. Administering this antibody in preclinical models reduced EMT, metastasis, and chemotherapy resistance. Early clinical trials confirmed its ability to decrease EMT in patient tumor biopsies, marking a significant advance in directly targeting the EMT process [98].
Experimental Models and Methodologies
In Vivo Zebrafish Screening Protocol
The following protocol leverages the optical clarity and genetic tractability of zebrafish to screen for anti-metastasis compounds [94].
- Materials: Wild-type or transgenic zebrafish lines, chemical libraries (e.g., FDA-approved drugs), dimethyl sulfoxide (DMSO), embryo medium, stereomicroscope.
- Procedure:
- Embryo Collection & Preparation: Collect zebrafish embryos and maintain at 28.5°C. At the 2-cell stage, dechorionate embryos if necessary.
- Chemical Exposure: At the sphere stage (4 hours post-fertilization, hpf), array embryos into multi-well plates. Expose experimental groups to test chemicals dissolved in vehicle (e.g., 0.1% DMSO). Include vehicle-only control groups.
- Incubation & Fixation: Incubate embryos until control embryos reach the 90% epiboly stage (9 hpf). This is a 5-hour exposure window.
- Phenotypic Analysis: Under a stereomicroscope, score each embryo for epiboly progression. Compare treated embryos to controls. "Hit" chemicals are defined as those causing a significant delay or arrest in epiboly.
- Validation: Positive hits must be validated in secondary assays, such as mammalian cell-based EMT assays or mouse models of metastasis.
In Vitro EMT and Metastasis Assays
A suite of in vitro tools is available to dissect specific steps of the metastatic cascade [70].
- Classical Migration/Invasion Assays:
- Transwell Migration/Invasion Assay: Cells are seeded in a transwell insert. For invasion assays, the membrane is coated with Matrigel. Cells that migrate/invade through the pores to the lower chamber are quantified after staining.
- Scratch/Wound Healing Assay: A confluent cell monolayer is scratched, and the rate of gap closure is measured over time, indicating collective cell migration.
- 3D Culture Models:
- Spheroids and Organoids: These models better recapitulate the tumor architecture and are used to study local invasion into a surrounding extracellular matrix (ECM).
- Proteolytic Activity Assays:
- The urokinase plasminogen activator (uPA) and its receptor (uPAR) system, along with Matrix Metalloproteinases (MMPs) like MMP-2 and MMP-9, are critical for ECM degradation and intravasation. Their activity can be measured using zymography or ELISA [70].
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Research Reagents and Models for Investigating EMT and Metastasis
| Tool/Reagent | Function/Application | Example Use |
|---|---|---|
| Zebrafish Embryo Model | Phenotypic, high-throughput chemical screening based on conserved gastrulation movements [94]. | Screening FDA-approved drug libraries for epiboly interruption [94]. |
| Foxa2/Sox17 Reporter mESCs | Time-resolved lineage tracing of endoderm formation via epithelial plasticity, distinct from classical EMT [26]. | Studying Foxa2's role as an EMT suppressor during definitive endoderm specification [26]. |
| ZO-1-GFP Reporter | Live imaging of tight junctions and apical surface dynamics during cell ingression [6]. | Quantifying ratchet-like apical constriction in mouse epiblast cells [6]. |
| Anti-Netrin-1 Antibody | Therapeutic agent to block Netrin-1/UNC5B signaling, inhibiting EMT [98]. | Preclinical validation of EMT inhibition in xenograft models and patient-derived tissues [98]. |
| uPA/uPAR System Assays | Quantifying proteolytic activity essential for ECM degradation and intravasation [70]. | Correlating uPAR levels with disseminated tumor cell fate and metastatic potential [70]. |
Signaling Pathways and Conceptual Workflows
The following diagrams illustrate the core signaling nexus and an integrated discovery pipeline derived from gastrulation research.
The study of gastrulation and the primitive streak has evolved from a purely embryological pursuit to a critical source of insights for oncology. The dissection of EMT and the discovery of epithelial plasticity mechanisms reveal the profound cellular reprogramming that powers both development and metastasis. The quantitative genetic parallels and the success of therapies like Netrin-1 inhibition provide a compelling validation of this approach. Future research must continue to leverage sophisticated in vivo and in vitro models to further unravel the nuances of these shared pathways, with the ultimate goal of translating this foundational knowledge into novel, effective strategies to combat metastatic cancer and overcome therapy resistance.
Metabolic reprogramming represents a fundamental adaptive process in both physiological development and pathological cancer progression. This whitepaper examines the intricate metabolic alterations occurring during epithelial-to-mesenchymal transition (EMT), with particular emphasis on primitive streak formation as a developmental paradigm that mirrors cancer metastasis. We demonstrate how core metabolic pathways—including glycolysis, glutaminolysis, and lipid metabolism—are co-opted across these contexts to support biosynthetic demands, energy production, and cellular plasticity. Through comprehensive analysis of experimental data and emerging therapeutic approaches, this review provides researchers with methodological frameworks for investigating metabolic reprogramming and identifies potential targets for intervention in cancer and developmental disorders.
Metabolic reprogramming describes the dynamic alterations in cellular metabolic pathways that enable adaptation to changing physiological demands or environmental conditions. This process has gained recognition as a critical hallmark in both normal development and cancer progression [99] [100]. During embryogenesis, tightly regulated metabolic shifts provide the energy and biosynthetic precursors necessary for rapid cell proliferation, differentiation, and tissue patterning. Similarly, cancer cells hijack these evolved metabolic strategies to support uncontrolled growth, survival in nutrient-poor environments, invasion, and metastasis [99] [101].
The epithelial-to-mesenchymal transition (EMT) serves as a paradigm for understanding how metabolic reprogramming facilitates cellular plasticity in both contexts. EMT describes the process wherein epithelial cells lose cell-cell adhesion and polarity while acquiring mesenchymal characteristics including enhanced migratory capacity, invasiveness, and resistance to apoptosis [102] [103]. In development, EMT is essential for primitive streak formation, gastrulation, and neural crest delamination. In cancer, EMT drives tumor dissemination, stemness, and therapy resistance [104] [103]. Emerging evidence demonstrates that successful execution of EMT requires substantial metabolic rewiring to meet the bioenergetic and biosynthetic demands of this phenotypic transformation [102] [105] [101].
This review examines the parallel metabolic reprogramming events in developmental and cancer-associated EMT, with particular focus on primitive streak formation as a key developmental model. We provide comprehensive analysis of core altered pathways, detailed experimental methodologies, and therapeutic implications for targeting metabolic vulnerabilities in cancer.
Metabolic Pathways in Development and Cancer
Glycolytic Reprogramming
The Warburg effect or aerobic glycolysis, characterized by preferential conversion of glucose to lactate even under oxygen-rich conditions, was first observed in cancer cells but also occurs during developmental processes [99] [106]. In both contexts, glycolytic shifting supports rapid biomass accumulation and redox homeostasis.
Table 1: Glycolytic Reprogramming in Development and Cancer
| Component | Developmental Role | Cancer Association | Functional Significance |
|---|---|---|---|
| GLUT1/3 | Nutrient uptake in pre-implantation embryos [106] | Upregulated in multiple cancers [99] | Enhanced glucose import |
| HK2 | Embryonic tissue development | TGF-β-induced EMT in mammary cells [102] | Commits glucose to glycolysis |
| PFKFB3 | Developmental angiogenesis | TGF-β-induced EMT [102] | Controls glycolytic rate via F2,6BP |
| LDHA | Blastocyst formation [106] | Upregulated in tumors [99] | Regenerates NAD+ for glycolysis |
| PKM2 | Proliferating embryonic cells | TGF-β-induced EMT in colon cancer [104] | Directs flux to biomass synthesis |
During primitive streak formation, embryonic cells undergo EMT and exhibit increased glycolytic flux similar to cancer cells. This metabolic adaptation provides ATP rapidly while generating glycolytic intermediates for biosynthetic pathways, including the pentose phosphate pathway for nucleotide synthesis and glycerol-3-phosphate for lipid membrane production [106] [101]. The transcription factor HIF-1α, stabilized in both hypoxic tumor microenvironments and during normal development, coordinates this glycolytic shift by transactivating glycolytic genes and directly promoting EMT through induction of Snail, Twist, and other EMT-transcription factors (EMT-TFs) [101].
Mitochondrial Metabolism and Glutaminolysis
While glycolysis often predominates, mitochondrial metabolism remains essential in both developmental and cancer contexts. The tricarboxylic acid (TCA) cycle not only generates ATP through oxidative phosphorylation but also supplies critical intermediates for biosynthesis [99] [105].
Table 2: Mitochondrial Metabolic Alterations in EMT
| Metabolite/Enzyme | Change During EMT | Functional Consequence |
|---|---|---|
| Citrate | Increased [102] | Lipid membrane synthesis |
| Succinate/Fumarate | Marked increase [102] | TCA cycle anaplerosis; potential signaling roles |
| Malate | Significant increase [102] | Aspartate production for nucleotide synthesis |
| Glutaminolysis | Enhanced [102] [105] | TCA cycle replenishment; nitrogen source |
| IDH/FH/SDH mutations | Associated with EMT [104] | Oncometabolite accumulation; epigenetic changes |
Glutaminolysis emerges as a critical pathway in both developmental and cancer EMT. Glutamine serves as a nitrogen donor for nucleotide and amino acid synthesis while its carbon skeleton (α-ketoglutarate) replenishes TCA cycle intermediates (anaplerosis) [99] [105]. In TGF-β-induced EMT, glutaminase (GLS1) expression increases, rendering cells dependent on glutamine metabolism [104]. Similarly, during embryonic development, rapidly proliferating cells exhibit heightened glutamine consumption to support biosynthetic demands.
Lipid Metabolism
Lipid metabolic reprogramming represents another shared feature. During EMT, both cancer and developing cells alter lipid metabolism to support membrane biogenesis for rapidly dividing cells and energy storage.
Table 3: Lipid Metabolic Changes in EMT
| Pathway/Enzyme | Alteration in EMT | Biological Impact |
|---|---|---|
| Fatty Acid Synthesis | Context-dependent regulation [104] | Membrane biogenesis |
| Fatty Acid Oxidation | Increased in some models [105] | Alternative energy source |
| Choline Metabolism | TGF-β induces phosphocholine production [102] | Phospholipid membrane synthesis |
| Acetyl-CoA | Increased mitochondrial production [106] | Histone acetylation; epigenetic regulation |
The metabolic intermediate acetyl-CoA serves as a crucial node integrating energy metabolism, lipid synthesis, and epigenetic regulation. Glucose-derived acetyl-CoA contributes to histone acetylation, modifying chromatin accessibility and transcriptional programs during both development and EMT [106].
Experimental Approaches and Methodologies
Metabolomic Profiling Techniques
Nuclear Magnetic Resonance (NMR) spectroscopy and mass spectrometry represent cornerstone technologies for quantifying metabolic changes during EMT.
NMR Spectroscopy Protocol (adapted from [102])
- Cell Culture: NMuMG mouse mammary epithelial cells treated with TGF-β (2-5 ng/mL) for 12-48 hours to induce EMT
- Metabolite Extraction: Use methanol:chloroform:water (2:1:1) extraction at -20°C
- Sample Preparation: Resuspend dried extracts in deuterated solvent (D₂O) with 0.025% TSP as chemical shift reference
- Data Acquisition: Employ 1D ¹H NMR with presaturation for water suppression; NOESY-presat pulse sequence; 256 scans; 6.5 μs 90° pulse; 2.5 s acquisition time; 1.0 s relaxation delay
- Data Analysis: Process with NMR software (e.g., Chenomx, MNova); reference to TSP (0.0 ppm); quantify metabolites using electronic reference library
Mass Spectrometry-Based Metabolomics
- Liquid Chromatography-MS: Reverse-phase chromatography for hydrophobic metabolites; HILIC for polar metabolites
- Stable Isotope Tracing: Use ¹³C-glucose or ¹³C-glutamine to track metabolic flux through pathways
- Data Processing: XCMS, MZmine for peak picking; MetaboAnalyst for statistical analysis
Genome-Scale Metabolic Modeling
Constraint-Based Reconstruction and Analysis (COBRA) provides a computational framework for integrating omics data with metabolic networks to predict flux states.
Workflow for EMT Metabolic Modeling [105]
- Network Reconstruction: Use human metabolic reconstruction (e.g., Recon3D)
- Transcriptomic/Proteomic Integration: Map time-course EMT data to metabolic genes using algorithms like iMAT
- Flux Prediction: Apply parsimonious Flux Balance Analysis (pFBA) to estimate metabolic fluxes
- Sensitivity Analysis: Perform in silico reaction knockouts to identify metabolic vulnerabilities
- Validation: Compare predictions with experimental data (e.g., CRISPR screens, metabolite measurements)
Figure 1: Genome-scale metabolic modeling workflow for identifying EMT metabolic dependencies
Functional Metabolic Assays
Seahorse Extracellular Flux Analysis
- Glycolytic Function: Measure extracellular acidification rate (ECAR) after sequential injection of glucose, oligomycin, and 2-DG
- Mitochondrial Respiration: Measure oxygen consumption rate (OCR) after sequential injection of oligomycin, FCCP, and rotenone/antimycin A
- EMT Conditions: Compare epithelial vs. mesenchymal states following TGF-β induction
Isotope Tracing Experiments
- Glucose Tracing: Use U-¹³C-glucose to track label incorporation into lactate, TCA intermediates, and other metabolites
- Glutamine Tracing: Use U-¹³C-glutamine to assess glutaminolytic flux and TCA cycle activity
- Sample Processing: Quench metabolism with cold methanol; analyze by LC-MS or GC-MS
The Scientist's Toolkit: Essential Research Reagents
Table 4: Key Reagents for Investigating Metabolic Reprogramming in EMT
| Reagent/Category | Specific Examples | Research Application | Key References |
|---|---|---|---|
| EMT Inducers | TGF-β (2-5 ng/mL); TNF-α; Hypoxia (1-2% O₂) | Induce mesenchymal transition | [102] [104] |
| Metabolic Inhibitors | 2-DG (glycolysis); CB-839 (GLS1); AZD3965 (MCT1) | Pathway inhibition studies | [100] |
| Isotope Tracers | U-¹³C-glucose; U-¹³C-glutamine; ¹³C-palmitate | Metabolic flux analysis | [102] [105] |
| Kinase Inhibitors | RSM-93A (CHKα inhibitor) | Target choline metabolism in EMT | [102] |
| Antibodies | E-cadherin; N-cadherin; Vimentin; Snail | EMT marker validation | [102] [104] |
Metabolic Regulation of EMT in Primitive Streak Formation
During embryonic development, the primitive streak forms through coordinated EMT events that enable the generation of mesoderm and endoderm lineages. This developmental EMT shares remarkable metabolic similarities with cancer-associated EMT.
Glycolytic Activation: Cells undergoing EMT at the primitive streak exhibit heightened glucose uptake and lactate production, mirroring the Warburg effect observed in cancer cells [106]. This glycolytic shift supports rapid proliferation and biomass accumulation while maintaining redox balance through lactate export.
Glutamine Dependence: Similar to observations in cancer models [105], developing embryos utilize glutamine as both an energy substrate and nitrogen donor. Glutaminolysis provides α-ketoglutarate for TCA cycle anaplerosis, supporting aspartate and nucleotide synthesis essential for rapid cell division.
Metabolite-Mediated Epigenetic Regulation: Metabolic intermediates including acetyl-CoA, S-adenosylmethionine (SAM), and α-ketoglutarate function as cofactors or substrates for epigenetic enzymes, linking metabolic state to chromatin organization and transcriptional programs [106] [107]. This metabolic-epigenetic axis coordinates the expression of EMT-transcription factors during both primitive streak formation and cancer progression.
Figure 2: Metabolic regulation of EMT through epigenetic mechanisms
Therapeutic Targeting of Metabolic Reprogramming
The shared metabolic vulnerabilities between developmental and cancer EMT present attractive therapeutic opportunities. Several classes of metabolic inhibitors are under investigation:
Glycolytic Inhibitors
- 2-Deoxyglucose (2-DG): Competitive hexokinase inhibitor; limits glycolytic flux
- Lonidamine: Targets mitochondrial-bound hexokinase
- FX11: LDHA inhibitor; blocks lactate production
Glutaminolysis Targeting
- CB-839: Potent glutaminase (GLS1) inhibitor; currently in clinical trials for solid tumors [100]
- V-9302: Competitive antagonist of ASCT2 glutamine transporter
Other Metabolic Targets
- MEDI9447: Anti-CD73 antibody; inhibits adenosine-mediated immunosuppression [100]
- Enasidenib/Ivosidenib: Mutant IDH2/IDH1 inhibitors; FDA-approved for AML [100]
- RSM-93A: Choline kinase α inhibitor; mitigates TGF-β-induced EMT [102]
Clinical development of metabolic therapies faces challenges including patient stratification, on-target toxicities to normal proliferating cells, and metabolic plasticity enabling resistance. Combination approaches targeting complementary pathways or coupling metabolic inhibitors with conventional chemotherapy may yield improved outcomes.
Metabolic reprogramming represents a conserved adaptive mechanism supporting the dynamic cellular transitions required for both embryonic development and cancer progression. The striking parallels between metabolic alterations during primitive streak formation and cancer EMT highlight fundamental biological principles of cellular plasticity. Understanding these shared metabolic networks provides not only insight into normal development but also reveals targetable vulnerabilities in cancer. Future research directions should include single-cell metabolomic approaches to characterize metabolic heterogeneity in EMT intermediate states, development of more specific metabolic inhibitors with reduced toxicity, and exploration of metabolic immunomodulation in the tumor microenvironment. The continued dissection of metabolic reprogramming in development and cancer will undoubtedly yield novel therapeutic paradigms for targeting metastatic disease.
The epithelial-mesenchymal transition (EMT) represents a critical plasticity mechanism that drives cancer metastasis, therapy resistance, and disease recurrence. This whitepaper provides a comprehensive technical evaluation of therapeutic strategies targeting EMT pathways in clinical oncology. We systematically analyze the molecular basis of EMT, assess current clinical trial outcomes, and detail experimental methodologies for investigating EMT inhibition. Within the context of primitive streak research, we examine how developmental EMT programs are co-opted in malignancy and how this informs therapeutic targeting. Despite promising preclinical data demonstrating that EMT pathway inhibition can restore treatment sensitivity and suppress metastatic progression, clinical translation faces significant challenges including pathway redundancy, adaptive resistance, and the dynamic plasticity of partial EMT states. This review synthesizes current evidence to guide researchers and drug development professionals in advancing targeted interventions against EMT-driven malignancy.
Epithelial-mesenchymal transition (EMT) is an evolutionarily conserved process fundamental to metazoan development, first described by Elizabeth Hay in studies of amphibian limb regeneration [45]. During mammalian gastrulation, EMT enables the formation of the primitive streak, through which epiblast cells ingress to give rise to the mesoderm and definitive endoderm [4] [10]. This developmental program is characterized by the dissolution of epithelial characteristics—including cell-cell adhesion and apicobasal polarity—and acquisition of mesenchymal traits that confer migratory capacity [45] [8].
In oncogenesis, cancer cells reactivate this developmental program, utilizing EMT to drive invasion, metastasis, and therapy resistance [4] [108]. The reverse process, mesenchymal-epithelial transition (MET), facilitates metastatic colonization at distant sites [109]. EMT is rarely complete in cancer; instead, cells often stabilize in hybrid epithelial/mesenchymal (E/M) states that exhibit enhanced plasticity and adaptive capabilities [108] [110]. This plasticity represents a fundamental challenge for therapeutic targeting, as the dynamic nature of these states enables rapid adaptation to therapeutic pressure.
The molecular regulation of EMT involves complex signaling networks including TGF-β, Wnt/β-catenin, Notch, ERK, and PI3K/AKT pathways [45] [108] [111]. These pathways converge on core transcription factors (Snail, Slug, Twist, ZEB1/2) that orchestrate the EMT program by repressing epithelial markers (e.g., E-cadherin) and activating mesenchymal markers (e.g., vimentin, N-cadherin, fibronectin) [45] [109]. Understanding the reactivation of this primitive streak-associated program in cancer provides the foundation for developing targeted therapies against EMT-driven malignancy.
Molecular Mechanisms of EMT: Signaling Pathways and Therapeutic Targets
Core Signaling Pathways Driving EMT
Table 1: Key Signaling Pathways in EMT and Their Therapeutic Targeting
| Pathway | Key Components | EMT-Inducing Mechanisms | Therapeutic Inhibitors |
|---|---|---|---|
| TGF-β | TGF-βR, Smad2/3/4, Smad7 | Induces Snail/Slug; Represses E-cadherin; Activates MAPK | Galunisertib (TGF-βRI inhibitor) |
| Wnt/β-catenin | Frizzled, LRP5/6, GSK-3β, β-catenin | Stabilizes β-catenin; Enhances transcription of EMT-TFs | PRI-724 (β-catenin/CBP inhibitor) |
| Notch | Notch receptor, DLL/Jagged, NICD, CSL | Upregulates Snail, Twist, Slug; Represses E-cadherin | Dibenzazepine (DBZ), DAPT (γ-secretase inhibitors) |
| RTK/Ras/MAPK | EGFR, HER2, Ras, Raf, MEK, ERK | Activates Twist, Snail; Cross-talks with TGF-β signaling | Trametinib (MEK inhibitor), Dasatinib (SRC inhibitor) |
| PI3K/AKT | PI3K, PDK1, AKT, mTOR | Phosphorylates EMT-TFs; Enhances stability and nuclear localization | MK-2206 (AKT inhibitor) |
The TGF-β pathway serves as a primary inducer of EMT across multiple carcinoma types [108]. TGF-β ligand binding initiates canonical signaling through Smad2/3 phosphorylation and complex formation with Smad4, leading to nuclear translocation and transcriptional activation of EMT master regulators including Snail and Twist [108]. Concurrently, non-canonical TGF-β signaling activates MAPK and PI3K/AKT pathways, creating reinforcing loops that stabilize the mesenchymal state [108]. The Wnt/β-catenin pathway promotes EMT through cytoplasmic stabilization and nuclear translocation of β-catenin, where it associates with TCF/LEF transcription factors to activate mesenchymal genes [45] [109]. Notch signaling contributes to EMT through γ-secretase-mediated release of the Notch intracellular domain (NICD), which translocates to the nucleus and forms a complex with CSL to activate Snail, Twist, and Slug expression while repressing E-cadherin [108] [111].
EMT Transcription Factors as Central Regulators
The core EMT transcription factors (EMT-TFs) function as master regulators of the phenotype switch. Snail and Slug (members of the SNAIL family) bind E-box elements in the CDH1 promoter, recruiting repressor complexes that silence E-cadherin expression [109]. ZEB1 and ZEB2 similarly repress epithelial genes while activating mesenchymal programs, with their activity modulated by regulatory miRNAs from the miR-200 family [109] [110]. Twist proteins heterodimerize with E-proteins to regulate target gene expression, with non-acetylated Twist recruiting NuRD repressor complexes to epithelial genes, while diacetylated Twist activates mesenchymal genes through BRD8 interaction [109]. These EMT-TFs exhibit significant functional redundancy yet maintain context-specific roles across cancer types, creating challenges for selective therapeutic targeting.
Figure 1: Core Signaling Pathways Regulating EMT. Multiple signaling cascades converge on master transcription factors that coordinately repress epithelial genes and activate mesenchymal programs, driving the invasive phenotype.
Clinical Trial Evaluation of EMT-Targeted Therapies
Clinical Trial Design and Endpoints
Clinical evaluation of EMT inhibitors presents unique methodological challenges, particularly in endpoint selection and biomarker validation. Unlike cytotoxic therapies that primarily target tumor shrinkage, EMT-directed agents aim to suppress invasion, prevent metastasis, and reverse therapy resistance—outcomes that may not immediately translate into conventional response metrics [112]. Current trials increasingly incorporate correlative biomarker studies assessing EMT marker shifts (e.g., E-cadherin restoration, vimentin reduction) in paired tumor biopsies [109] [112]. Circulating tumor cells (CTCs) with hybrid E/M phenotypes are emerging as promising non-invasive biomarkers for monitoring EMT inhibition and disease progression [109].
Representative Clinical Trial Data
Table 2: Clinical Trial of EMT Pathway Inhibitors in Solid Tumors
| Trial Identifier | Cancer Type | Intervention | Target | Primary Endpoints | Key Findings | Reference |
|---|---|---|---|---|---|---|
| NCT01990196 | Unfavorable-risk Prostate Cancer | ADT ± Trametinib ± Dasatinib | MEK/SRC | EMT markers (N-cadherin, vimentin) by IHC | No significant difference in EMT markers between arms; MAP2K1/2 and SRC RNA decreased in all arms | [112] |
| Preclinical | Glioblastoma | 20(S)-Rg3, Quercetin, Honokiol | Multiple pathways | Invasion, migration, stemness | Suppressed EMT; Enhanced TMZ sensitivity; Reduced invasion | [111] |
| Multiple | Colorectal Cancer | Natural compounds (Curcumin, Resveratrol) | TGF-β, Wnt, NF-κB | Metastasis prevention, Chemosensitization | Reversed EMT; Attenuated invasion; Restored drug sensitivity | [109] [110] |
A phase 2 randomized trial in unfavorable-risk prostate cancer (NCT01990196) exemplifies the challenges in clinical targeting of EMT [112]. This study evaluated neoadjuvant androgen deprivation therapy (ADT) alone or combined with the MEK inhibitor trametinib or SRC inhibitor dasatinib. The primary endpoint assessed EMT marker expression (N-cadherin and vimentin) following treatment. Contrary to preclinical hypotheses, neither trametinib nor dasatinib provided significant additional suppression of EMT markers compared to ADT alone [112]. All treatment arms showed reduced MAP2K1, MAP2K2, and SRC RNA abundance, suggesting that ADT itself may exert inhibitory effects on these pathways. This trial highlights the complex regulation of EMT in clinical settings and potential redundancy in pathway targeting.
Experimental Protocols for EMT Inhibition Studies
In Vitro Assessment of EMT Inhibition
Protocol: Evaluation of Compound Efficacy Against TGF-β-Induced EMT
- Cell Culture and EMT Induction: Plate epithelial cancer cells (e.g., prostate PC-3, breast MDA-MB-468) in 6-well plates at 50,000 cells/well. After 24 hours, replace media with serum-free medium containing TGF-β (2-5 ng/mL) with or without the investigational compound at varying concentrations.
- Morphological Analysis: Capture phase-contrast images at 24, 48, and 72 hours post-treatment. Quantify the percentage of cells with mesenchymal (elongated, spindle-shaped) morphology versus epithelial (cobblestone) appearance.
- Immunofluorescence Staining: At 72 hours, fix cells with 4% PFA, permeabilize with 0.1% Triton X-100, and block with 5% BSA. Incubate with primary antibodies against E-cadherin, vimentin, and N-cadherin overnight at 4°C, followed by appropriate fluorescent secondary antibodies. Image using confocal microscopy and quantify fluorescence intensity.
- Western Blot Analysis: Harvest cells in RIPA buffer, separate proteins by SDS-PAGE, transfer to PVDF membranes, and probe with antibodies against E-cadherin, N-cadherin, vimentin, Snail, Slug, and Twist. Use GAPDH or β-actin as loading controls.
- Transwell Invasion Assay: Seed 50,000 serum-starved cells in Matrigel-coated transwell inserts with test compounds in the upper chamber. Place complete growth medium in the lower chamber as chemoattractant. After 24 hours, fix, stain with crystal violet, and count invaded cells.
In Vivo Metastasis Models
Protocol: Experimental Metastasis Assay with EMT Inhibition
- Animal Model: Utilize 6-8 week old immunocompromised mice (e.g., NOD/SCID or NSG).
- Cell Labeling and Injection: Label cancer cells with luciferase for bioluminescence imaging. Pre-treat cells with EMT inhibitor or vehicle control for 72 hours. Inject 1×10^6 cells via tail vein (for lung metastasis model) or intrasplenically (for liver metastasis model).
- Treatment Protocol: Administer EMT inhibitor or vehicle control via oral gavage or intraperitoneal injection beginning one day post-injection and continuing daily.
- Metastasis Monitoring: Image weekly using in vivo bioluminescence imaging. Quantify metastatic burden by photon flux in regions of interest.
- Endpoint Analysis: Euthanize mice at predetermined endpoint or when signs of distress appear. Harvest lungs, liver, and other organs for ex vivo imaging, histology, and immunohistochemistry analysis of EMT markers.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for EMT Studies
| Reagent Category | Specific Examples | Application in EMT Research | Technical Notes |
|---|---|---|---|
| EMT Inducers | Recombinant TGF-β, TNF-α, EGF | Induction of EMT in epithelial cell lines | TGF-β at 2-5 ng/mL optimal for most cell lines; pre-test time course |
| Pathway Inhibitors | SB431542 (TGF-βRI), XAV939 (Wnt), DAPT (Notch) | Pathway-specific EMT inhibition | Use concentration gradients; monitor cytotoxicity |
| Antibodies for IHC/IF | Anti-E-cadherin, Anti-vimentin, Anti-N-cadherin | EMT marker quantification in cells and tissues | Validate specificity with knockout controls |
| EMT Transcription Factor Antibodies | Anti-Snail, Anti-Slug, Anti-Twist, Anti-ZEB1 | Detection of EMT-TFs in nuclear extracts | Requires nuclear fractionation for Western blot |
| qPCR Assays | CDH1, VIM, CDH2, SNAI1, TWIST1 | mRNA expression profiling of EMT markers | Normalize to multiple housekeeping genes |
| Invasion Assay Materials | Matrigel, Transwell inserts, Boyden chambers | Quantification of invasive capacity | Optimize Matrigel concentration for each cell type |
| 3D Culture Systems | Basement membrane extract, Collagen I matrices | Study EMT in physiologically relevant context | Allows observation of hybrid E/M states |
Challenges and Future Perspectives in EMT-Targeted Therapy
The clinical translation of EMT-targeted therapies faces several formidable challenges. First, the profound plasticity and heterogeneity of EMT states enable rapid adaptive resistance to single-agent pathway inhibition [4] [108]. Second, the developmental origins of EMT create on-target toxicity concerns, as physiological EMT processes remain active in tissue repair and wound healing [45]. Third, current clinical trial endpoints may be insufficiently sensitive to detect meaningful biological effects of EMT inhibition, particularly in the adjuvant setting where prevention of metastasis is the primary goal [112].
Future directions should focus on several key areas: (1) Rational combination therapies that simultaneously target multiple EMT pathways or co-target EMT and complementary resistance mechanisms; (2) Advanced biomarker strategies incorporating liquid biopsy platforms to monitor dynamic EMT plasticity in response to therapy; (3) Nanotechnology approaches to enhance blood-brain barrier penetration for EMT inhibition in glioblastoma and other CNS malignancies [111]; (4) Immunotherapy combinations leveraging the emerging understanding of EMT-immune system interactions; (5) Natural product exploration to identify novel EMT-inhibitory scaffolds with unique mechanisms of action [110].
The continued elucidation of EMT regulation in primitive streak formation and embryonic development will provide critical insights for therapeutic targeting in cancer [26] [8]. As we deepen our understanding of the evolutionary conservation and context-specific adaptations of this fundamental developmental program, we move closer to effective therapeutic strategies for suppressing EMT-driven malignancy and improving outcomes in advanced cancer patients.
Conclusion
The EMT of the primitive streak is not merely a historical developmental event but a living process whose principles are subverted in human disease, particularly cancer. Understanding the conserved signaling pathways, transcriptional networks, and cellular behaviors that govern this primary EMT provides an indispensable framework for deciphering the mechanisms of tumor metastasis, fibrosis, and therapy resistance. Future research must leverage advanced models and technologies to further dissect the dynamics of partial EMT states and their specific contributions to pathology. The translational potential is vast; by targeting the reactivated embryonic programs in cancer cells, novel therapeutic strategies can be devised to inhibit metastasis and overcome treatment failure, ultimately bridging the fundamental biology of the embryo to clinical oncology.
References
- 1. Primitive streak [https://en.wikipedia.org/wiki/Primitive_streak]
- 2. Epithelial-mesenchymal transitions: the importance of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2689100/]
- 3. Guidelines and definitions for research on epithelial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7250738/]
- 4. Epithelial-mesenchymal transition [https://www.sciencedirect.com/science/article/abs/pii/S0092867425010256]
- 5. Epithelial to Mesenchymal Transition History [https://www.mdpi.com/2218-273X/11/6/782]
- 6. A ratchet-like apical constriction drives cell ingression ... [https://elifesciences.org/articles/84019]
- 7. Stem cell-based human embryo models [https://pmc.ncbi.nlm.nih.gov/articles/PMC12395930/]
- 8. Epithelial-Mesenchymal Transition Drives Three ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2021.639244/full]
- 9. Guidelines and definitions for research on epithelial ... [https://www.nature.com/articles/s41580-020-0237-9]
- 10. Mouse primitive streak forms in situ by initiation of epithelial to ... [https://pubmed.ncbi.nlm.nih.gov/22170865/]
- 11. The Epithelial-to-Mesenchymal Transition (EMT) in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8189433/]
- 12. BMP signaling and EMT during and at the end of gastrulation [https://pmc.ncbi.nlm.nih.gov/articles/PMC2958622/]
- 13. The basics of epithelial-mesenchymal transition - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC2689101/]
- 14. Cellular dynamics of EMT: lessons from live in vivo imaging of ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-021-00761-8]
- 15. Epithelial-mesenchymal transitions: insights from ... [https://pubmed.ncbi.nlm.nih.gov/22949611/]
- 16. FGF/FGFR signaling in health and disease [https://www.nature.com/articles/s41392-020-00222-7]
- 17. β-Catenin Regulates Primitive Streak Induction through ... [https://www.sciencedirect.com/science/article/pii/S1934590915001198]
- 18. Vg1-Nodal heterodimers are the endogenous inducers of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5745085/]
- 19. A role for Vg1/Nodal signaling in specification of the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3621495/]
- 20. FGF Signaling Pathway: A Key Regulator of Stem Cell ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7040165/]
- 21. Roles of FGF signaling in stem cell self-renewal ... [https://www.aging-us.com/article/100369/text]
- 22. The role of Wnt/β-catenin-lin28a/let-7 axis in embryo ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-020-00562-5]
- 23. Wnt signaling induces epithelial–mesenchymal transition ... [https://www.nature.com/articles/labinvest2011201]
- 24. Role of epithelial-mesenchymal transition factor SNAI1 and its targets in ovarian cancer aggressiveness [https://www.oaepublish.com/articles/2394-4722.2023.34]
- 25. Role of epithelial-mesenchymal transition factor SNAI1 and its targets in ovarian cancer aggressiveness - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10673625/]
- 26. Epithelial cell plasticity drives endoderm formation during ... [https://www.nature.com/articles/s41556-021-00694-x]
- 27. SNAI1 promotes epithelial-mesenchymal transition and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11657537/]
- 28. SNAI1-Mediated Epithelial-Mesenchymal Transition Confers ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0066558]
- 29. Molecular mechanisms of TWIST1‐regulated transcription ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10626429/]
- 30. TWIST1 is a critical downstream target of the HGF/MET ... [https://pubmed.ncbi.nlm.nih.gov/38485737/]
- 31. Twist1-induced epithelial-mesenchymal transition ... [https://www.oncotarget.com/article/10974/text/]
- 32. Signaling mechanisms of the epithelial-mesenchymal transition [https://pmc.ncbi.nlm.nih.gov/articles/PMC4372086/]
- 33. Primitive Streak - an overview [https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/primitive-streak]
- 34. Two distinct epithelial-to-mesenchymal transition programs ... [https://www.nature.com/articles/s43018-024-00839-5]
- 35. Epithelial-Mesenchymal Transition Drives Three ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7905045/]
- 36. Changes in cellular composition shape the inductive ... [https://www.nature.com/articles/s41467-025-63154-2]
- 37. Transitions between epithelial and mesenchymal states and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2958623/]
- 38. Evidence for Partial Epithelial-to-Mesenchymal Transition ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3128786/]
- 39. In vivo visualization and characterization of epithelial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4873431/]
- 40. 3D gastruloids: a novel frontier in stem cell-based in vitro ... [https://pubmed.ncbi.nlm.nih.gov/34304959/]
- 41. A continuous totipotent-like cell-based embryo model ... [https://www.nature.com/articles/s41556-025-01793-9]
- 42. Self-patterning of human stem cells into post-implantation ... [https://www.nature.com/articles/s41586-023-06354-4]
- 43. A 3D in vitro model to explore the inter-conversion between ... [https://www.nature.com/articles/srep27072]
- 44. An In Vitro Model That Recapitulates the Epithelial to ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0017083]
- 45. Epithelial–Mesenchymal Transition in Cancer: Insights Into ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12394892/]
- 46. Molecular mechanism involved in epithelial to ... [https://www.sciencedirect.com/science/article/abs/pii/S0003986121002332]
- 47. Identification of prognostic biomarkers related to epithelial ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2025.1600546/full]
- 48. Modeling the dynamics of EMT reveals genes associated ... [https://www.nature.com/articles/s41540-025-00512-2]
- 49. Proteome Landscape of Epithelial-to-Mesenchymal ... [https://www.sciencedirect.com/science/article/pii/S1535947621001031]
- 50. Mechanistic insights into promotion of non-small cell lung ... [https://pubmed.ncbi.nlm.nih.gov/40787462/]
- 51. Synthetic augmentation of cancer cell line multi-omic ... [https://www.nature.com/articles/s41467-024-54771-4]
- 52. Cellular dynamics of EMT: lessons from live in vivo imaging of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8296657/]
- 53. Long-term live imaging, cell identification and cell tracking ... [https://elifesciences.org/articles/107534]
- 54. Quantifying cell migration in real time [https://www.news-medical.net/whitepaper/20250827/Quantifying-cell-migration-in-real-time-Methods-and-applications.aspx]
- 55. Epithelial-to-mesenchymal transition proceeds through ... [https://elifesciences.org/articles/74866]
- 56. Epithelial-to-Mesenchymal Transition in Fibrosis: Concepts ... [https://www.frontiersin.org/journals/pharmacology/articles/10.3389/fphar.2021.737570/full]
- 57. Perturbomics: CRISPR–Cas screening-based functional ... [https://www.nature.com/articles/s12276-025-01487-0]
- 58. CRISPR-mediated modeling and functional validation of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6955235/]
- 59. Genome-wide CRISPR screens define determinants of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10348683/]
- 60. Off-target effects in CRISPR-Cas genome editing for ... [https://www.sciencedirect.com/science/article/pii/S2162253125001908]
- 61. Bioengineering strategies to control epithelial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8068500/]
- 62. Epithelial-to-mesenchymal transition and different ... [https://www.sciencedirect.com/science/article/abs/pii/S0955067420300648]
- 63. Emerging Concepts of Hybrid Epithelial-to-Mesenchymal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7697085/]
- 64. Epithelial/mesenchymal plasticity: how have quantitative ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5496493/]
- 65. Epithelial-mesenchymal transitions: the importance of ... [https://www.jci.org/articles/view/38019]
- 66. Hybrid EMT and inflammation: the deadly alliance [https://www.sciencedirect.com/science/article/pii/S2667394025000292]
- 67. Pan-cancer quantitation of epithelial-mesenchymal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8832824/]
- 68. Epithelial-to-mesenchymal transition (EMT) and cancer ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12144846/]
- 69. Epithelial-to-Mesenchymal Transition in the Light of ... [https://pubmed.ncbi.nlm.nih.gov/34072345/]
- 70. Epithelial-to-mesenchymal transition (EMT) and cancer ... [https://molecular-cancer.biomedcentral.com/articles/10.1186/s12943-025-02338-2]
- 71. Dependence of cell fate potential and cadherin switching ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11838492/]
- 72. Targeting epithelial-mesenchymal transition signaling ... [https://pubmed.ncbi.nlm.nih.gov/39959483/]
- 73. Epithelial-mesenchymal transition orchestrates tumor ... [https://translational-medicine.biomedcentral.com/articles/10.1186/s12967-025-06422-5]
- 74. CAV1 promotes epithelial-to-mesenchymal transition (EMT ... [https://www.frontiersin.org/journals/immunology/articles/10.3389/fimmu.2025.1523855/full]
- 75. Molecular Mechanisms Regulating Epithelial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12072817/]
- 76. JCI - Biomarkers for epithelial - mesenchymal transitions [https://www.jci.org/articles/view/36183]
- 77. EMT markers | Abcam [https://www.abcam.com/en-us/technical-resources/research-areas/marker-guides/epithelial-to-mesenchymal-transition-markers]
- 78. Significance of E - cadherin and Vimentin as epithelial - mesenchymal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7355153/]
- 79. Detection of Biomarkers for Epithelial-Mesenchymal ... [https://www.imrpress.com/journal/FBL/27/4/10.31083/j.fbl2704127/htm]
- 80. Signaling networks guiding epithelial–mesenchymal transitions during... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11158989/]
- 81. The mouse primitive streak forms in situ by initiation of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3266444/]
- 82. What is the epithelial to mesenchymal transition? - Mechanobiology... [https://www.mbi.nus.edu.sg/mbinfo/what-is-the-epithelial-to-mesenchymal-transition/]
- 83. Comparative Study of Transcriptomics-Based Scoring ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7100584/]
- 84. Pan-cancer quantitation of epithelial-mesenchymal transition ... [https://translational-medicine.biomedcentral.com/articles/10.1186/s12967-021-03227-0]
- 85. Protocol for inferring epithelial-to-mesenchymal transition ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10789639/]
- 86. and definitions for research on epithelial-mesenchymal... Guidelines [https://pubmed.ncbi.nlm.nih.gov/32300252/]
- 87. The basics of epithelial-mesenchymal transition [https://www.jci.org/articles/view/39104]
- 88. The molecular mechanisms and therapeutic strategies of EMT ... [https://jhoonline.biomedcentral.com/articles/10.1186/s13045-022-01347-8]
- 89. Innovative Approaches to EMT-Related Biomarker ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12467340/]
- 90. Embryonic mesenchyme, mesenchymal tumors and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12618774/]
- 91. Cell-context response to germ layer differentiation signals ... [https://www.nature.com/articles/s41467-025-60348-6]
- 92. Epithelial-to-mesenchymal transition drives cancer genomic ... [https://jeccr.biomedcentral.com/articles/10.1186/s13046-025-03402-x]
- 93. Gastrulation Research Reveals Novel Details About ... [https://www.helmholtz-munich.de/en/idr/news-detail/gastrulation-research-reveals-novel-details-about-embryonic-development]
- 94. Gastrulation Screening to Identify Anti-metastasis Drugs in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9548519/]
- 95. Role of imbalanced gut microbiota in promoting CRC ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-024-01615-9]
- 96. The Link Between the Gut Microbiome and Bone Metastasis [https://pmc.ncbi.nlm.nih.gov/articles/PMC11593804/]
- 97. Delineating the nexus between gut-intratumoral ... [https://www.sciencedirect.com/science/article/pii/S2352187224000767]
- 98. A new drug prevents EMT, metastasis and resistance to ... [https://ecancer.org/en/news/23461-a-new-drug-prevents-emt-metastasis-and-resistance-to-anti-cancer-therapy]
- 99. Cancer metabolic reprogramming: importance, main features ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3969803/]
- 100. Metabolic Reprogramming and Epithelial-Mesenchymal Plasticity: Opportunities and Challenges for Cancer Therapy - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC7252305/]
- 101. Targeting the Metabolic Reprogramming That Controls ... [https://www.frontiersin.org/journals/oncology/articles/10.3389/fonc.2017.00040/full]
- 102. Metabolic Reprogramming of Mammary Epithelial Cells during TGF-β-Induced Epithelial-to-Mesenchymal Transition - PMC [https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8464788/]
- 103. Reprogramming during epithelial to mesenchymal transition under the control of TGFβ - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4594534/]
- 104. Metabolic reprogramming and epithelial-to-mesenchymal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6049610/]
- 105. Genome-scale modeling identifies dynamic metabolic ... [https://www.nature.com/articles/s42003-024-07408-7]
- 106. Metabolic Reprogramming of Stem Cell Epigenetics - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4672395/]
- 107. Epigenetic regulation in metabolic diseases: mechanisms ... [https://www.nature.com/articles/s41392-023-01333-7]
- 108. The impact of epithelial-mesenchymal transition (EMT ... [https://biosignaling.biomedcentral.com/articles/10.1186/s12964-024-01957-4]
- 109. Epithelial-mesenchymal transition in colorectal cancer ... [https://www.nature.com/articles/s41420-025-02593-8]
- 110. Can Natural Products Targeting EMT Serve as the Future ... [https://www.mdpi.com/1420-3049/27/22/7668]
- 111. Molecular drivers of epithelial-mesenchymal transition ... [https://www.sciencedirect.com/science/article/abs/pii/S0344033825003048]
- 112. A Phase 2 Randomized Clinical Trial [https://www.sciencedirect.com/science/article/abs/pii/S2588931125002871]