Spatial Transcriptomics in Embryonic Development: From Foundational Principles to Advanced Applications
This article provides a comprehensive overview of cutting-edge methods for detecting mRNA spatial expression in embryos, a rapidly advancing field crucial for understanding development and disease.
Spatial Transcriptomics in Embryonic Development: From Foundational Principles to Advanced Applications
Abstract
This article provides a comprehensive overview of cutting-edge methods for detecting mRNA spatial expression in embryos, a rapidly advancing field crucial for understanding development and disease. We explore the foundational principles of spatial transcriptomics, compare the performance and applications of major methodological platforms like Visium, Stereo-seq, and MERFISH, and address key troubleshooting and optimization challenges. Furthermore, we detail strategies for validating spatial data and integrating it with single-cell RNA sequencing. Designed for researchers, scientists, and drug development professionals, this review synthesizes current benchmarks and offers practical guidance for selecting and implementing these powerful technologies in embryonic research.
The Blueprint of Life: Why Spatial mRNA Localization is Fundamental to Embryogenesis
Defining Spatial Transcriptomics and Its Critical Role in Developmental Biology
Spatial transcriptomics is a cutting-edge suite of techniques that maps gene expression data within the context of a tissue, uncovering how different genes are expressed in specific locations and thereby revealing how cells are arranged, interact, and function in their native environments [1]. For decades, understanding the genetic programs of embryonic development relied on bulk RNA-sequencing, which analyzes the averaged gene expression of a population of cells, or single-cell RNA-sequencing (scRNA-seq), which identifies individual cells and their distinct features but requires tissue dissociation, irrevocably losing all spatial information [1] [2]. Developmental biology is inherently spatial; the formation of organs and complex structures is driven by precise, location-specific gene expression. Spatial transcriptomics overcomes historical limitations by preserving this crucial anatomical context, enabling researchers to directly investigate the spatiotemporal regulation of life processes during development [2]. This whitepaper defines the core methodologies of spatial transcriptomics, details its experimental protocols, and highlights its transformative application in deciphering the molecular architecture of embryonic development.
Core Methodologies and Technical Principles
Spatial transcriptomics technologies can be broadly classified into two main categories based on their underlying principles for capturing spatial gene expression: imaging-based methods and sequencing-based methods [3] [4].
Imaging-Based Technologies
Imaging-based technologies employ single-molecule fluorescence in situ hybridization (smFISH) or in situ sequencing (ISS) to visualize RNA molecules directly within cells or tissues.
- smFISH and Derivatives: Techniques like MERFISH, seqFISH+, and RNAscope use fluorescently labeled probes that hybridize to target transcripts. They achieve high multiplexing through sequential rounds of hybridization, imaging, and probe stripping, generating unique fluorescent signatures or binary barcodes for each gene [2] [3] [4].
- In Situ Sequencing (ISS): Methods such as STARmap and FISSEQ involve reverse transcribing mRNA to cDNA within the tissue, followed by in situ amplification and sequencing-by-ligation. This allows for the direct reading of nucleotide sequences in their original tissue location [2] [4].
Sequencing-Based Technologies
Sequencing-based technologies integrate spatially barcoded arrays with next-generation sequencing.
- Spatially Barcoded Arrays: Platforms like 10x Visium and Stereo-seq use slides coated with oligonucleotides containing spatial barcodes. When a tissue section is placed on the slide, mRNA is captured, and the spatial barcode is incorporated during cDNA synthesis. Subsequent sequencing allows transcripts to be mapped back to their original locations [2] [3].
- Laser Capture Microdissection (LCM): An earlier approach, LCM uses a laser to precisely isolate cells from specific histological regions under microscopic guidance, after which the captured material is subjected to transcriptomic analysis [2] [4].
Table 1: Comparison of Major Commercial Spatial Transcriptomics Platforms
| Platform | Technology Category | Spatial Resolution | Detection Sensitivity | Throughput (Number of Genes) | Key Differentiating Feature |
|---|---|---|---|---|---|
| 10x Visium [3] | Sequencing-based | 55 μm spots | High | Whole transcriptome | Cost-effective, user-friendly workflow for broad discovery |
| 10x Visium HD [3] | Sequencing-based | 2 μm bins | High | Whole transcriptome | Enhanced resolution from smaller, bin-based features |
| Xenium [3] | Imaging-based | Single-cell & subcellular | High | ~500-1,000 genes | Hybrid ISS/ISH chemistry with padlock probes & RCA |
| CosMx SMI [3] [5] | Imaging-based | Single-cell & subcellular (<50 nm) | High | Up to 19,000 transcripts | Combinatorial color & position optical signature |
| MERSCOPE [3] | Imaging-based | Single-cell & subcellular | High | ~500-1,000 genes | Binary barcoding strategy for error correction |
| Stereo-seq [2] [3] | Sequencing-based | Subcellular (0.5 μm DNB) | High | Whole transcriptome | Ultra-high resolution via DNA Nanoball (DNB) arrays |
| GeoMx DSP [3] [5] | Sequencing-based | Region of Interest (ROI) | High | Whole transcriptome | UV-cleavage of tags from ROIs selected by morphology |
Diagram 1: A hierarchical classification of core Spatial Transcriptomics methodologies, distinguishing between imaging-based and sequencing-based foundational technologies.
Experimental Protocols: From Tissue to Data
A typical spatial transcriptomics workflow involves several critical steps, from sample preparation to data analysis. The following protocol, integrating steps from platforms like Visium and imaging-based methods, provides a generalized framework for a spatially-resolved gene expression experiment.
Tissue Preparation and Sectioning
- Sample Collection and Preservation: Tissues of interest (e.g., embryonic mouse brain) are collected. The choice of preservation is critical: fresh-frozen tissue is optimal for maximum RNA integrity, while Formalin-Fixed Paraffin-Embedded (FFPE) tissue preserves morphological structure and is compatible with some platforms (e.g., Visium V2 with CytAssist) but presents challenges due to RNA cross-linking and degradation [1] [3].
- Cryosectioning: Frozen tissues are embedded in Optimal Cutting Temperature (OCT) compound and sectioned into thin slices (typically 5-20 μm thick) using a cryostat. Sections are then carefully transferred onto specific spatial transcriptomics slides (e.g., Visium slides) [6].
On-Slide Library Construction
This stage varies significantly between sequencing-based and imaging-based platforms.
For Sequencing-Based Platforms (e.g., 10x Visium):
- Staining and Imaging: The tissue section on the slide is stained with Hematoxylin and Eosin (H&E) and imaged using a brightfield microscope to obtain a high-resolution histological reference image [6].
- Permeabilization: The tissue is treated with a permeabilization enzyme to puncture cell membranes, allowing mRNA molecules to diffuse out and reach the capture probes on the slide. The permeabilization time must be optimized for each tissue type to balance mRNA capture efficiency and spatial resolution [1] [3].
- mRNA Capture and Reverse Transcription: The released mRNA binds to the spatially barcoded oligo-dT probes on the slide. Reverse transcription occurs on the slide, creating cDNA molecules that incorporate the spatial barcode and a Unique Molecular Identifier (UMI) [3].
- Second-Strand Synthesis and Library Preparation: The cDNA is released from the slide, and the second strand is synthesized. The library is then prepared for sequencing via PCR amplification, adding sequencing adapters [3].
For Imaging-Based Platforms (e.g., CosMx, Xenium):
- Fixation and Permeabilization: The tissue section is fixed and permeabilized to allow access for probes.
- Probe Hybridization: A panel of gene-specific probes is hybridized to the target mRNAs within the tissue. For example, CosMx uses primary probes with readout domains, while Xenium uses padlock probes that are circularized upon target binding [3].
- Signal Amplification and Readout: Signal is amplified enzymatically (e.g., via Rolling Circle Amplification in Xenium) or via branched fluorophores (in CosMx). The slide is then imaged using a high-resolution fluorescence microscope [3].
- Cyclic Imaging: For multiplexing, the process involves multiple rounds of fluorescence imaging, followed by chemical stripping of the fluorophores or cleavage of the signal, and re-hybridization with new probes to decode different genes in each cycle [3] [4].
Sequencing, Data Processing, and Analysis
- Sequencing and Base Calling: For sequencing-based methods, the libraries are run on a next-generation sequencer (e.g., Illumina). For imaging-based methods, the data consists of multi-round TIFF images [3].
- Spatial Coordinate Alignment: The gene expression data (count matrix) is aligned with the histological image based on the spatial barcodes (sequencing-based) or through image analysis (imaging-based) to reconstruct the spatial map [6] [7].
- Downstream Bioinformatics Analysis: This includes spot/cell clustering, differential expression analysis, cell-type deconvolution using scRNA-seq reference data, and trajectory inference to model developmental processes [6] [7].
Diagram 2: A consolidated experimental workflow for Spatial Transcriptomics, showing parallel paths for sequencing-based and imaging-based platforms converging on integrated data analysis.
The Scientist's Toolkit: Essential Reagents and Materials
Table 2: Key Research Reagent Solutions for Spatial Transcriptomics
| Item | Function | Example Use Case |
|---|---|---|
| Visium Spatial Gene Expression Slide [3] | Glass slide pre-printed with millions of spatially barcoded oligos for mRNA capture. | Foundation for 10x Visium workflow; captures whole transcriptome from a tissue section. |
| CytAssist Instrument [3] | Compact instrument that transfers probes from a standard glass slide to a Visium slide. | Enables spatial analysis of FFPE tissues in the Visium V2 workflow. |
| Gene-Specific Probe Panels (e.g., for CosMx, Xenium) [3] [5] | Pre-designed pools of DNA probes that bind to target mRNA sequences for imaging-based detection. | Hypothesis-driven targeting of specific gene sets in development (e.g., patterning genes). |
| Permeabilization Enzyme [1] [3] | Enzyme (e.g., protease) that treats tissue sections to allow mRNA to diffuse out of cells. | Critical step optimized for each tissue type (e.g., embryonic brain, muscle) to maximize yield. |
| Fluorophore-Labeled Readout Probes [3] [4] | Fluorescently tagged secondary probes that bind to primary probes for signal generation. | Used in cyclic FISH methods (MERFISH, CosMx) to read out gene-specific barcodes. |
| Padlock Probes [3] | Single-stranded DNA probes that circularize upon hybridization to a cDNA target for RCA. | Key reagent in Xenium and other in situ sequencing platforms for specific amplification. |
| DNA Nanoballs (DNBs) [3] | Rolling circle amplification products of barcoded oligos that form a high-density capture array. | The core of the Stereo-seq platform, providing subcellular spatial resolution. |
Application in Developmental Biology: Decoding the Embryo
Spatial transcriptomics has become indispensable for dissecting cell-fate decisions and unraveling mechanisms of tissue morphogenesis [2]. A seminal application is the construction of high-resolution spatiotemporal atlases of embryonic development.
A 2025 study on the late-stage embryonic and postnatal mouse brain exemplifies this power [6]. Researchers used the 10x Visium platform to profile coronal sections of the forebrain at embryonic day 17 (E17) and postnatal day 0 (P0). Integrated analysis with adult mouse brain data allowed them to identify spatially coherent gene expression profiles across 25 distinct clusters corresponding to anatomical regions like the neocortex, thalamus, and hippocampus.
Crucially, the study uncovered novel molecular markers specific to developmental stages and structures. For instance, through differential expression analysis, genes Folr1 (folate receptor 1) and Car12 (carbonic anhydrase 12) were identified as specific markers for the choroid plexus, a key structure in cerebrospinal fluid production. Validation by in situ hybridization (ISH) confirmed their exclusive presence in the choroid plexus at E17 and P0, unlike the known marker Msx1, which was also expressed in the leptomeninges [6]. This demonstrates the technology's ability to discover more precise markers for defining embryonic structures.
Furthermore, the study identified a novel marker, Etl4, for the dorsal endopiriform nucleus (DEn) within the complex embryonic claustrum/DEn structure. This finding, complemented by integration with single-cell data, revealed an internal substructure of heterogeneous cell types, providing unprecedented insight into the molecular architecture of the developing brain [6].
Beyond the brain, spatial transcriptomics is being applied to other developing systems. In skeletal muscle research, it has been used to explore the cellular crosstalk essential for regeneration, a process that recapitulates aspects of development. It has helped characterize spatially defined interactions between muscle stem cells and inflammatory or resident cells, which are dysregulated in degenerative diseases like Duchenne Muscular Dystrophy (DMD) [4].
Future Perspectives and Challenges
The field of spatial transcriptomics is rapidly evolving, yet it faces several challenges that must be addressed to maximize its potential. Technical limitations include the high cost of some platforms, the slow throughput depending on the number of genes analyzed, and the difficulty in achieving true single-cell resolution with some early methods, especially in densely packed tissues [1]. Sample preparation remains a hurdle, as precise tissue handling is critical for high-quality data, and not all platforms are universally compatible with FFPE samples [1].
The computational burden of managing and analyzing the immense complexity of spatial datasets is significant [1] [7]. A 2025 review highlighted at least 24 different computational tools dedicated solely to the alignment and integration of multiple tissue slices, a non-trivial task due to tissue heterogeneity and plasticity [7]. Future progress hinges on robust automated methodologies for these tasks.
Despite these challenges, the future is promising. Innovations are continuously emerging, such as a new computational method from the Broad Institute that eliminates the need for intensive imaging by inferring spatial locations through molecular biology and algorithms, potentially making spatial genomics more accessible and scalable [8]. The market is also shifting towards integrated multi-omics solutions, with companies developing platforms that combine transcriptomics with proteomics and other data modalities from the same sample [9] [10]. The integration of Artificial Intelligence (AI) is poised to bridge the gap between routine pathology and deep molecular analysis, enabling predictive biology and transforming spatial biology into a cornerstone of clinical research and drug development in the coming decade [9] [10].
The precise spatiotemporal regulation of gene expression is the fundamental architect of embryonic development, orchestrating a complex sequence of cell fate specification, migration, and tissue morphogenesis. Spatial mRNA gradients serve as key molecular instruments in this symphony, conveying positional information that guides the formation of body axes and organ primordia. Traditional single-cell RNA-sequencing (scRNA-seq) has revolutionized cell type identification but sacrifices crucial spatial context, limiting our understanding of how transcriptional programs are organized within native tissue architecture [11]. The emergence of spatial transcriptomics (ST) technologies has bridged this critical gap, enabling genome-wide expression profiling while preserving spatial locality. These advances now permit the quantitative investigation of regionalized gene expression patterns within developing embryos, providing unprecedented insights into the molecular mechanisms governing early development [12] [13]. This technical guide synthesizes recent breakthroughs in spatial genomics, detailing how spatially resolved mRNA gradients establish embryonic patterning from axis formation to organ primordia specification, with particular emphasis on experimental methodologies and computational frameworks driving these discoveries.
Technological Foundations of Spatial Transcriptomics
Spatial transcriptomics encompasses a suite of technologies that enable transcriptome-wide gene expression analysis while maintaining spatial information within tissue sections. These methods can be broadly categorized into sequencing-based approaches (e.g., Slide-seq, Visium) that use DNA-barcoded spots on a surface to capture mRNA, and imaging-based approaches (e.g., MERFISH) that visualize individual RNA molecules through sequential hybridization and imaging [13]. The fundamental challenge in ST analysis involves aligning and integrating multiple tissue slices to reconstruct three-dimensional expression patterns, a task complicated by tissue heterogeneity, technical variability, and differences in experimental protocols [7].
Recent computational innovations have significantly advanced this field. For instance, sc3D was developed specifically to reconstruct and explore three-dimensional 'virtual embryos' from serial Slide-seq sections, enabling quantitative analysis of regionalized gene expression along developmental axes [12]. Similarly, SEU-3D was created to reconstruct full digital embryos at single-cell resolution during early organogenesis, allowing investigation of gene expression in native spatial context [14]. A groundbreaking method from the Broad Institute eliminates the need for specialized imaging equipment altogether by using computational approaches to reconstruct spatial locations of gene expression based on molecular diffusion between "transmitter" and "receiver" beads, dramatically reducing costs while enabling analysis of larger tissue areas [8].
Table 1: Key Spatial Transcriptomics Technologies for Embryonic Research
| Technology | Spatial Resolution | Throughput | Key Applications in Development | References |
|---|---|---|---|---|
| Slide-seq | 10 μm | Transcriptome-wide | Whole-embryo mapping at E8.5-E9.5; 3D virtual embryo reconstruction | [12] |
| MERFISH | Single-cell | Targeted gene panels (300+ genes) | Human fetal cortex layer specification; single-cell spatial atlas | [11] |
| 10x Visium | 55 μm (1-30 cells/spot) | Transcriptome-wide | Developing mouse brain; integration with scRNA-seq | [6] [7] |
| Computational Array Reconstruction | Subcellular | Transcriptome-wide | Large tissue sections (up to 7 cm); no specialized equipment required | [8] |
Spatial Patterning of Embryonic Axes
Neural Tube Patterning
The developing neural tube exhibits a remarkably precise stratification of gene expression along its dorsoventral (DV) axis, which is essential for subsequent neuronal subtype diversification. Spatial transcriptomic maps of mouse embryos at E8.5-E9.5 have enabled systematic identification of genes with restricted spatial expression along this axis. For example, sc3D-based reconstruction of whole E8.5 and E9.0 embryos revealed several previously unannotated genes with distinct spatial patterns in the neural tube [12]. The analysis quantified regionalization through a localization score that ranked genes based on their spatial restriction, identifying markers that delineate specific developmental axes and presumptive anatomical structures.
Brain Compartmentalization
Between E8.5 and E9.5, the anterior neural tube develops into three distinct vesicles (prosencephalon, mesencephalon, and rhombencephalon) that form the primordial brain. High-resolution spatial transcriptomics demonstrates that spatial restriction of gene expression precedes anatomical segregation. Genes including Foxg1 (confined to rostral prosencephalon), Barhl2 (expressed caudally marking presumptive diencephalon), and Rax (defining the future optic cup) exhibit confined expression patterns before clear anatomical boundaries form [12]. Unsupervised spatial RNA velocity analysis without prior knowledge of cell states reveals distinct trajectory dynamics, with converging or diverging patterns corresponding to stepwise transitions or cellular steady states. These analyses specifically highlight boundary regions like the mesencephalon-rhombencephalon boundary, marked by restrictive expression of Otx2 in the mesencephalon and Gbx2 in the rhombencephalon [12].
Figure 1: Gene Expression Patterns During Brain Compartmentalization. Spatial transcriptomics reveals restricted expression of key markers before anatomical boundaries form.
Organ Primordia Specification
Cardiac Primordium Formation
Early organogenesis represents a crucial stage characterized by extensive cell fate specification to initiate organ formation. At E7.75 in mouse embryos, researchers have characterized a primordium determination zone (PDZ) that forms along the anterior embryonic-extraembryonic interface [14]. Through single-cell 3D transcriptome analysis and space-informed gene-cell co-embedding approaches, studies have elucidated how coordinated signaling communications across germ layers contribute to cardiac primordium formation. The PDZ represents a specialized microenvironment where precise spatial organization of signaling molecules and transcription factors establishes the foundation for heart development, ultimately leading to the formation of the cardiac crescent [14].
Cortical Layer Specification in the Brain
The human cerebral cortex develops a complex six-layer structure that emerges much earlier than previously recognized. Spatial transcriptomic analysis of human fetal cortex using MERFISH (multiplexed error-robust fluorescence in situ hybridization) revealed that excitatory neuron subtypes exhibit distinct laminar distributions in the cortical plate as early as gestational week 15—approximately three months before the emergence of cytoarchitectural layers visible through conventional histology [11]. This study analyzed over 18 million single cells across eight cortical areas and seven developmental timepoints, demonstrating that molecular layer specification precedes morphological differentiation. To quantitatively analyze cell localization, researchers developed a framework assigning each cell a relative height (RH) representing its normalized laminar position between apical and basal surfaces, enabling precise mapping of cortical depth (CD) for excitatory neuron subtypes [11].
Enteric Nervous System Patterning
The enteric nervous system (ENS) displays a complex 3D architecture along the gut axis, with specific transcriptional states needed to control gut homeostasis. Whole-gut spatial genomic analysis (SGA) in zebrafish larvae has identified spatially variable genes (including hoxb5b, hoxa4a, etv1, and ret) that are regionalized along gut axes, suggesting precise spatiotemporal control of ENS development [15]. This patterning occurs during the differentiation of enteric neural progenitor cells (ENPs) as they migrate and differentiate into functionally diverse neuron types. The application of SGA to ENS development has provided new insights into its cellular transcriptional networks and interactions, establishing a baseline dataset for understanding gut neurodevelopmental disorders such as Hirschsprung disease [15].
Table 2: Key Organ Primordia and Their Spatial mRNA Gradients
| Organ Primordium | Key Spatial Patterning Genes | Developmental Stage | Technical Approach | Functional Significance | |
|---|---|---|---|---|---|
| Cardiac Crescent | Cross-germ-layer signaling networks | E7.75 mouse | Single-cell 3D transcriptomics (SEU-3D) | Establishes primordium determination zone (PDZ) | [14] |
| Cortical Layers | Excitatory neuron subtype markers | GW15 human | MERFISH + deep-learning segmentation | Molecular layering precedes cytoarchitecture | [11] |
| Enteric Nervous System | hoxb5b, hoxa4a, etv1, ret | 4-7 dpf zebrafish | Whole-mount HCR + 3D segmentation | Regional patterning along gut axes | [15] |
| Neural Tube | Otx2, Gbx2, Fgf8 (boundaries) | E8.5-E9.5 mouse | Slide-seq + sc3D reconstruction | Establishes brain compartment boundaries | [12] |
Experimental Methodologies and Protocols
Slide-seq for Whole-Embryo Spatial Transcriptomics
Slide-seq provides a powerful method for generating transcriptome-wide gene expression data at 10-μm spatial resolution, enabling reconstruction of whole embryonic structures [12]. The protocol involves:
Bead Array Preparation: DNA-barcoded beads with known spatial coordinates are arranged on a slide surface. Each bead contains unique spatial barcodes that serve as positional references.
Tissue Sectioning and Transfer: Embryos are cryosectioned at 10-μm thickness (sagittal sections for whole embryos). Sections are transferred onto the bead array, and tissue is permeabilized to release mRNA.
mRNA Capture and Library Preparation: Released mRNA molecules bind to proximal barcoded beads through poly-dT capture. After reverse transcription, sequencing libraries are constructed with unique molecular identifiers (UMIs) to quantify transcript abundance.
Sequencing and Data Processing: Libraries are sequenced on high-throughput platforms. The resulting data are processed to generate spatial expression matrices, with each bead linked to its original spatial coordinates and gene expression profile.
For the mouse embryo study, this approach recovered 533,116 high-quality beads with a median of 1,798 transcripts and 1,224 genes per bead, enabling comprehensive mapping of embryonic structures [12].
Computational 3D Reconstruction with sc3D
The sc3D computational pipeline transforms 2D spatial transcriptomic data into 3D "virtual embryos" through the following steps:
Section Alignment: Individual spatial transcriptomic arrays from serial sections are aligned along the z-axis using morphological features and marker genes as landmarks.
3D Grid Reconstruction: Aligned sections are interpolated to reconstruct a continuous 3D volume, with each voxel containing gene expression information.
Virtual In Situ Hybridization (vISH): The reconstruction enables quantitative analysis of gene expression gradients along any embryonic axis, inclination plane, or rotation angle.
Spatial Analysis: Genome-wide correlations between tissue volumes and expressing cell densities generate localization scores that rank genes based on spatial restriction [12].
This framework has demonstrated robustness to increasing inter-slice intervals with minimal distortion in rotation axes, enabling reliable 3D reconstruction from sparse section sampling [12].
MERFISH for Single-Cell Spatial Atlas of Human Fetal Cortex
MERFISH enables spatially resolved single-cell transcriptomics through sequential hybridization and imaging:
Gene Panel Design: Curate a panel of target genes (e.g., 300 genes including canonical markers and cluster-specific genes from scRNA-seq references).
Tissue Processing and Hybridization: Human fetal cortex sections are fixed and permeabilized. Gene-specific encoding probes with readout sequences are hybridized to target mRNAs.
Sequential Imaging and Stripping: Multiple rounds of fluorescence readout probe hybridization, imaging, and probe stripping sequentially decode the spatial positions of individual RNA molecules.
Single-Cell Segmentation: A custom deep-learning model based on CellPose 2.0 automates single-nucleus segmentation from nucleus-stained images. A modest dilation of nuclei-based cell masks enriches transcript counts without compromising cell identity precision [11].
This approach analyzed over 18 million single cells while maintaining true single-cell resolution, despite the high cell density of the fetal brain [11].
Figure 2: Experimental Workflow for Spatial Transcriptomics in Embryonic Development. The pipeline progresses from tissue preparation through computational reconstruction to visualization.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Essential Research Reagents for Spatial Transcriptomics of Embryonic Development
| Reagent/Resource | Specification | Function | Example Application | |
|---|---|---|---|---|
| DNA-barcoded Beads | 10-μm diameter, spatial barcodes | mRNA capture with spatial information | Slide-seq for whole-embryo mapping | [12] |
| MERFISH Encoding Probes | Gene-specific probes with readout sequences | Multiplexed mRNA detection through sequential hybridization | Single-cell spatial atlas of human fetal cortex | [11] |
| HCR Probes | Hybridization chain reaction amplifiers | Signal amplification for low-abundance transcripts | Whole-gut spatial genomic analysis in zebrafish | [15] |
| sc3D Software | Python package for 3D reconstruction | Alignment of serial sections into 3D virtual embryos | Quantitative analysis of gene expression gradients | [12] |
| CellPose 2.0 | Deep-learning nucleus segmentation | Automated single-cell identification in dense tissues | Human fetal cortex analysis at single-cell resolution | [11] |
| Uniform Manifold Approximation and Projection (UMAP) | Dimension reduction algorithm | Visualization of high-dimensional spatial data | Computational array reconstruction without imaging | [8] |
Spatial transcriptomics has fundamentally transformed our understanding of how mRNA gradients orchestrate embryonic development, from initial axis patterning to organ primordia specification. The technologies and methodologies reviewed here—including Slide-seq, MERFISH, and computational reconstruction approaches—provide powerful tools for constructing comprehensive spatiotemporal atlases of embryonic development at single-cell resolution. These advances have revealed that molecular patterning often precedes morphological changes, with gene expression gradients establishing developmental blueprints before physical structures emerge.
Future directions in the field will likely focus on increasing spatial resolution to subcellular levels, improving throughput for larger tissue sections, and enhancing multi-omics integration to simultaneously capture transcriptomic, proteomic, and epigenetic information within spatial context. Additionally, computational methods for data integration and analysis will continue to evolve, addressing current challenges in aligning heterogeneous tissue slices and reconstructing complex 3D architectures [7]. As these technologies become more accessible and cost-effective, they will undoubtedly uncover new principles of developmental biology and provide insights into the spatial dysregulation of gene expression underlying congenital disorders. The ongoing synthesis of spatial genomics with functional perturbation studies will further elucidate causal relationships between mRNA gradients and phenotypic outcomes, ultimately advancing both basic developmental biology and clinical applications in prenatal diagnosis and regenerative medicine.
A fundamental challenge in developmental biology is understanding how the precise spatial and temporal expression of genes directs the formation of a complex organism from a single cell. Recent technological advances have dramatically enhanced our ability to detect and quantify mRNA spatial expression in embryos, providing unprecedented insights into the molecular mechanisms of development. This whitepaper examines how three cornerstone model systems—zebrafish, mouse, and Xenopus—are being leveraged to dissect these intricate processes, with a focus on the innovative methodologies powering this research. These organisms provide complementary strengths for studying gene expression dynamics, enabling researchers to address questions spanning from basic developmental mechanisms to applications in toxicology and drug development.
Model System Strengths and Applications
Each model organism offers a unique set of advantages for developmental studies, allowing researchers to select the most appropriate system for their specific research questions.
Table: Key Characteristics of Model Embryonic Systems
| Organism | Key Strengths | Primary Research Applications | Genetic Tools Available |
|---|---|---|---|
| Zebrafish | Optical transparency, high fecundity, external development [16] | High-throughput chemical screening, live imaging of development [16], single-cell transcriptomics [17] | CRISPR/Cas9, transgenic reporters, metabolic labeling [18] [17] |
| Mouse | Close phylogenetic relation to humans, established embryology | Spatial transcriptomics, mechano-transcriptomics, pre-implantation development studies [19] [20] | Sophisticated transgenic models, CRISPR tools, spatial omics |
| Xenopus | Large embryos for micromanipulation, diploid genome (X. tropicalis), abundant maternal material | Proteomics, transcriptomics dynamics, high-throughput functional screening [21] [22] | CRISPRi, morpholinos, extensive RNA-seq databases [21] [22] |
Advanced Methodologies for mRNA Detection
Single-Cell Metabolic Labeling in Zebrafish
The combination of single-cell RNA-Seq (scRNA-Seq) with metabolic labeling represents a powerful approach for distinguishing newly-transcribed (zygotic) mRNA from pre-existing (maternal) mRNA in zebrafish embryogenesis [17]. This method involves injecting 4-thiouridine-triphosphate (4sUTP) into one-cell stage embryos, which is selectively incorporated into newly-transcribed RNA. After capturing mRNA on beads using Drop-Seq, a chemical conversion step creates T-to-C changes in sequencing reads, allowing precise quantification of zygotic transcription. Subsequent GRAND-SLAM analysis determines the fraction of newly-transcribed mRNA for each gene in individual cells, enabling researchers to quantify mRNA transcription and degradation rates at cell-type resolution during development [17].
Diagram: Workflow for Single-Cell Metabolic Labeling in Zebrafish Embryos
Spatial Mechano-Transcriptomics in Mouse
A novel computational framework enables the joint statistical analysis of transcriptional and mechanical signals in the context of spatial transcriptomics of developing mouse embryos [19]. This integrated mechano-transcriptomics approach begins with compiling input data including immunostained cell membranes, seqFISH images, and single-cell transcriptomic references. After processing and segmenting images to delineate cell boundaries, the pipeline employs a variational method of stress inference (VMSI) to quantify mechanical forces. This algorithm uses a nonplanar triangulation of junctional tensions to infer intracellular pressures and tensions at cell-cell junctions [19]. The resulting mechanical estimates are then statistically associated with transcriptomic readouts, accounting for spatial confounders to identify genes and ligand-receptor pairs associated with mechanical phenotypes.
Table: Quantitative mRNA Dynamics During Embryonic Development
| Biological Process | Organism | Key Finding | Quantitative Measurement |
|---|---|---|---|
| pri-mir-9 Expression | Zebrafish | Sequential activation of 7 pri-mir-9 loci creates sharp stepwise increase in mature miR-9 [18] | Late neuronal differentiation reduced by pri-mir-9-1 mutation [18] |
| Maternal-to-Zygotic Transition | Zebrafish | Metabolic labeling distinguishes maternal and zygotic transcripts at single-cell level [17] | Zygotic mRNA increases from 13% (4.3 hpf) to 41% (5.3 hpf) of cellular mRNA [17] |
| Pre-implantation Translation | Mouse | Polysome profiling reveals stage-specific delay in mRNA translation [20] | Identification of Eif1ad3 as essential translation factor at two-cell stage [20] |
| Proteome-MRNA Correlation | Xenopus | Mass action kinetics model links protein and mRNA dynamics [22] | Absolute levels of ~10,000 proteins and ~28,000 transcripts measured [22] |
CRISPR Interference (CRISPRi) in Xenopus
CRISPRi has emerged as an effective and suitable approach to suppress specific mRNA transcription in Xenopus tropicalis embryos [21]. The system utilizes a catalytically dead Cas9 (dCas9) fused to repressive domains such as KRAB-MeCP2 (dCas9-KM), which is guided to target genes by specific gRNAs to form a DNA recognition complex that interferes with transcription. Compared to CRISPR-Cas13 systems which showed limited efficacy in Xenopus, CRISPRi efficiently targeted both exogenous and endogenous transcripts [21]. Recent optimizations have identified that the KRAB domain from ZIM3 protein (ZIM3-KRAB) alone has comparable transcript targeting capacity to the traditional fusion repressor KRAB-MeCP2, providing an improved tool for exploring specific gene function during development [21].
Diagram: CRISPRi Mechanism for mRNA Knockdown in Xenopus
The Scientist's Toolkit: Essential Research Reagents
Table: Key Research Reagents for Embryonic mRNA Studies
| Reagent / Tool | Function | Example Application |
|---|---|---|
| 4sUTP (4-thiouridine-triphosphate) | Metabolic label incorporated into newly transcribed RNA | Distinguishing zygotic from maternal transcripts in zebrafish [17] |
| LNA (Locked Nucleic Acid) probes | High-affinity nucleic acid analogs for in situ detection | Detecting mature miR-9 expression in zebrafish embryos [18] |
| dCas9-KRAB-MeCP2 | CRISPR interference fusion protein for transcriptional repression | Specific mRNA knockdown in Xenopus tropicalis embryos [21] |
| Cyp3a65-GFP transgenic zebrafish | Reporter for metabolic enzyme expression | Assessing metabolic endocrine-disrupting chemicals in intestine [23] |
| seqFISH reagents | Sequential fluorescence in situ hybridization for spatial transcriptomics | Mapping mRNA expression with subcellular resolution in mouse embryos [19] |
| Polysome profiling reagents | Fractionation of ribosome-bound mRNA | Studying translational dynamics during mouse pre-implantation development [20] |
Applications in Drug Development and Toxicology
The zebrafish model has proven particularly valuable for high-throughput toxicological screening and mechanistic studies of chemical effects. A recent study leveraged zebrafish embryogenesis to evaluate the effects of 45 structurally diverse agrichemicals, linking transcriptome-wide changes to phenotypic outcomes [16]. This approach combined morphological assessment at 120 hours post-fertilization (hpf) with transcriptomic profiling at 48 hpf, prior to the onset of overt morphological effects. The study identified between 0 and 4,538 differentially expressed genes per chemical, with convergence on shared biological pathways including neurodevelopment and cytoskeletal organization [16]. This demonstrates how model systems can provide system-level insights for evaluating chemical toxicity and understanding how chemical structure influences biological activity during development.
The complementary strengths of zebrafish, mouse, and Xenopus model systems provide a powerful toolkit for dissecting the spatial and temporal dynamics of mRNA expression during embryonic development. Zebrafish offer unparalleled opportunities for live imaging and high-throughput screening, mouse provides the closest mammalian counterpart for human-relevant studies, and Xenopus enables sophisticated micromanipulation and high-resolution omics profiling. As the methods discussed—from single-cell metabolic labeling to spatial mechano-transcriptomics and advanced CRISPR perturbations—continue to evolve, they will further deepen our understanding of developmental gene regulation. These insights not only advance fundamental knowledge but also enhance our ability to assess chemical safety and develop novel therapeutic strategies, highlighting the enduring value of these model systems in biomedical research.
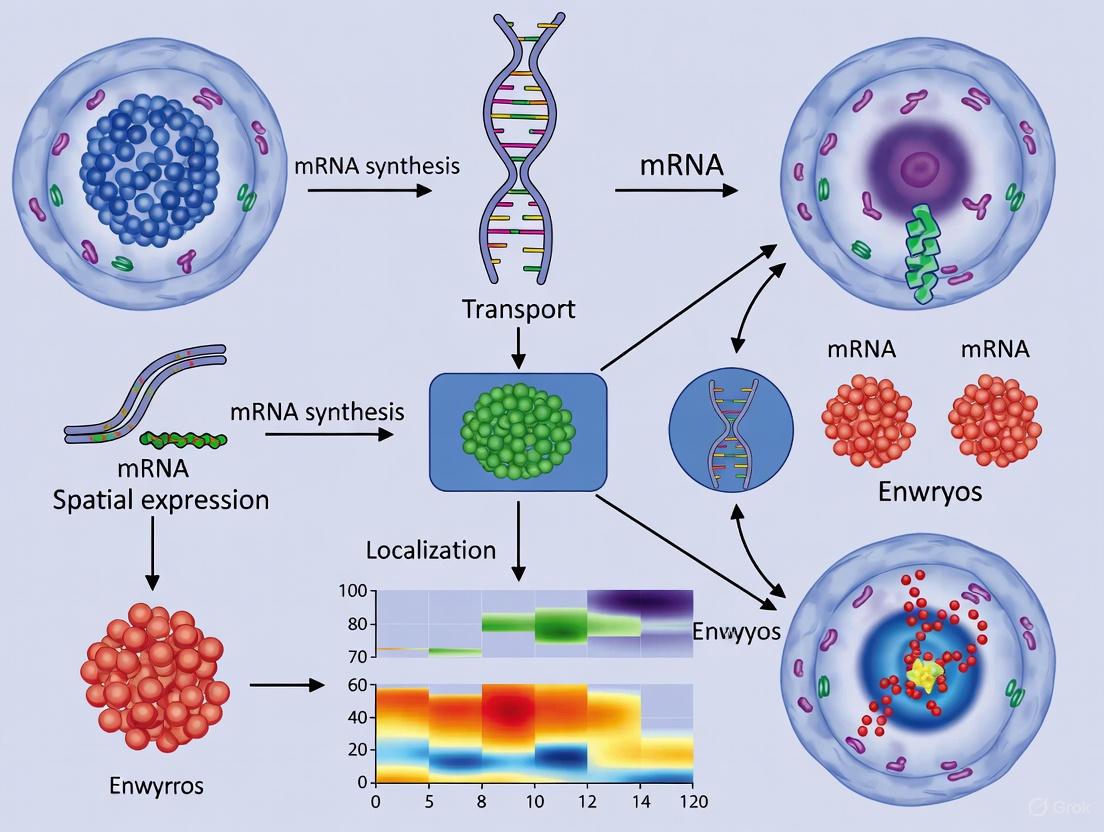
The precise spatial localization of mRNA expression within an embryo is not merely a consequence of development; it is a fundamental driver of the process. The emergence of spatial transcriptomics (ST) has revolutionized developmental biology by allowing researchers to visualize the intricate patterns of gene expression that direct cell fate, tissue patterning, and organ formation in situ. This whitepaper explores how these powerful technologies are unveiling the complex signaling pathways and cellular interactions that govern the development of the Enteric Nervous System (ENS) and early craniofacial structures. By preserving the anatomical context of gene expression, ST provides an unprecedented window into the spatiotemporal dynamics of embryogenesis, offering critical insights for understanding congenital disorders and advancing regenerative medicine strategies.
Case Study I: Enteric Nervous System Development and Hirschsprung's Disease
EMB in ENS Development: A Novel Genetic Regulator
The ENS, often described as a "second brain" in the gut, arises from enteric neural crest cells (ENCCs) that migrate, proliferate, and differentiate to form a complex network of neurons and glia essential for gastrointestinal motility, secretion, and blood flow [24] [25]. Defects in ENS development can lead to Hirschsprung's disease (HSCR), a congenital disorder characterized by the absence of ganglia in the distal colon, occurring in approximately 1 in 5,000 live births [24] [25].
Recent research has identified Embigin (EMB) as an evolutionarily conserved, essential regulator of ENS development. EMB is a transmembrane glycoprotein that facilitates ENS formation by recruiting the phosphatase complex PP2A to the cellular membrane. This recruitment reduces cytoplasmic dephosphorylation activity and promotes the activation of the PI3K-AKT signaling pathway, a critical pathway for cellular survival, proliferation, and migration [24].
Table 1: Key Findings from EMB Functional Studies
| Experimental Model | Phenotype of EMB Loss | Rescue Potential |
|---|---|---|
| Zebrafish | Decreased enteric neurons; impaired intestinal transit | Partial rescue with PI3K or AKT agonists |
| Mouse | HSCR-like aganglionosis | Not reported |
| In Vitro (gut explants/organoids) | Impaired ENCC proliferation and migration | Not reported |
Molecular Pathway and Experimental Validation
The mechanistic pathway of EMB action was delineated through a series of in vitro and in vivo experiments. In mouse gut explants and organoid cultures, the loss of EMB led to significant defects in both the proliferation and migration of ENCCs. Mechanistically, EMB binds to and recruits the protein phosphatase 2A (PP2A) complex to the cell membrane. This relocalization reduces the cytoplasmic pool of PP2A, thereby diminishing its dephosphorylation activity on key signaling molecules. This change in the phosphorylation landscape facilitates the activation of the PI3K-AKT pathway, a crucial driver of cell growth and movement. The functional importance of this pathway was confirmed when application of PI3K or AKT agonists partially restored ENS development in emb-mutant zebrafish [24]. Furthermore, sequencing of a human HSCR patient cohort revealed rare EMB variants, suggesting its potential contribution to the pathology of the disease in humans [24].
Case Study II: Craniofacial Development and Cell Fate Determination
High-Resolution Mapping of Palatogenesis
Craniofacial development serves as another powerful model for understanding spatiotemporal cell fate determination. A recent study employed a high-resolution seqFISH-based spatial transcriptomics platform to create a detailed spatiotemporal transcriptomic and cell-type atlas of cranial neural crest cell (CNCC)-derived mesenchymal lineage diversification during mouse palatogenesis [26]. This approach enabled the systematic definition of mesenchymal cell types by mapping their transcriptomic profiles directly to their spatial identities within the developing palate.
Integrative analysis of spatial transcriptomic data from embryonic days E12.5 to E15.5 revealed that mesenchymal lineages are established at or even prior to the initiation of palatogenesis. A key finding was the identification of a heterogeneous Sox9+ mesenchymal progenitor population at the onset of palatal development. Subpopulations within this group were already activating early lineage-specific markers, indicating an early commitment to distinct developmental paths [26].
Lineage Tracing Validation of Early Fate Decisions
The predictive power of the spatial transcriptomic data was confirmed through in vivo lineage tracing in transgenic mouse models. Using early lineage-specific markers identified through single-cell RNA sequencing (scRNA-seq) and spatial transcriptomics, researchers demonstrated that distinct mesenchymal populations are established as early as E10.5 to E11.5. This timeline precedes the physical formation of the palate, providing strong evidence that cell fate determination occurs well before the anatomical structures become apparent [26]. This work sheds light on the fundamental mechanisms of cell fate regulation during embryogenesis and has significant implications for understanding congenital anomalies like cleft palate.
Table 2: Spatiotemporal Analysis of Mouse Palatogenesis
| Research Component | Key Finding | Technical Method |
|---|---|---|
| Mesenchymal Progenitor | Identified a heterogeneous Sox9+ population at onset | scRNA-seq & seqFISH |
| Lineage Establishment | Major lineages determined at or before E12.5 | Integrated ST analysis (E12.5-E15.5) |
| Fate Commitment | Subpopulations express early lineage markers in vivo | Lineage tracing in transgenic mice |
| Developmental Timing | Fate determination occurs as early as E10.5-E11.5 | In vivo lineage tracing |
Advanced Methodologies for Spatial mRNA Detection
Cutting-Edge Spatial Transcriptomics Technologies
The case studies presented were enabled by advanced spatial transcriptomics technologies, which can be broadly classified into two categories [27]:
- Imaging-based Methods: These techniques visualize RNA molecules directly within tissues. Examples include MERFISH, seqFISH+, and STARmap. They rely on either fluorescent in situ hybridization (ISH) or in situ sequencing (ISS) and can achieve subcellular resolution. A key strength is their high resolution, but they typically require a predefined set of target genes.
- Sequencing-based Methods: Techniques like Slide-seq capture mRNA onto DNA-barcoded beads arrayed on a slide. After sequencing, computational methods map the gene expression back to the bead locations, allowing for genome-wide expression profiling without pre-selecting targets.
A recent innovation, computational array reconstruction, eliminates the need for intensive imaging in Slide-seq. Instead of imaging the bead array, it uses molecular diffusion between "transmitter" and "receiver" beads and a computational algorithm (UMAP) to reconstruct the original spatial locations of the beads. This approach significantly scales up the tissue area that can be analyzed and lowers the cost and equipment barriers [8].
Computational Integration and Subcellular Analysis
The integration of single-cell RNA sequencing (scRNA-seq) with spatial transcriptomics is crucial for achieving true single-cell resolution within a spatial context. Computational methods like SEU-TCA have been developed to map individual cells from scRNA-seq datasets onto spatial locations from ST data with high accuracy [28]. This integration is vital for elucidating highly dynamic cellular interactions during development.
Furthermore, as ST technologies achieve higher resolution, computational tools are evolving to analyze subcellular mRNA localization. ELLA is a statistical framework designed to model subcellular spatial variation of gene expression within cells. It uses an over-dispersed nonhomogeneous Poisson process to identify genes with specific subcellular localization patterns, which is essential for understanding mechanisms like localized protein synthesis and cell polarity [29].
The Scientist's Toolkit: Essential Reagents and Platforms
Table 3: Research Reagent Solutions for Spatial Transcriptomics and Lineage Analysis
| Reagent / Platform | Function / Application | Key Feature |
|---|---|---|
| Anti-EMB Antibody | Immunofluorescence staining to localize EMB protein in gut/colon tissues. | Validated for use in human and mouse tissues (e.g., Abcam ab127692) [24]. |
| seqFISH-based Platforms | High-resolution spatial mapping of gene expression in tissues like developing palate. | Allows for highly multiplexed, subcellular RNA detection [26]. |
| Slide-seq & Computational Array Reconstruction | Genome-wide spatial transcriptomics without pre-defined targets or specialized imaging. | Scalable, cost-effective analysis of large tissue areas (e.g., whole organs) [8]. |
| ELLA Software | Statistical detection of spatially variable genes at subcellular resolution. | Identifies mRNA localization patterns within cells from high-res ST data [29]. |
| SEU-TCA Algorithm | Integrates scRNA-seq and ST data to map single cells to spatial locations. | Superior deconvolution and spatial mapping accuracy for complex tissues [28]. |
| Wnt1Cre-R26RTomato Mouse Model | Lineage tracing and isolation of neural crest-derived cells for scRNA-seq. | Enables specific labeling and analysis of enteric neural crest cells [24]. |
The integration of sophisticated spatial transcriptomics technologies with precise functional experiments is fundamentally reshaping our understanding of embryonic development. The detailed examination of ENS development reveals how specific genes like EMB orchestrate critical processes such as ENCC migration and proliferation through defined signaling pathways, whose disruption leads to neurodevelopmental disorders like HSCR. Simultaneously, high-resolution spatiotemporal mapping of craniofacial development demonstrates that cell fate determination is an early and spatially pre-patterned event. These insights, powered by advanced computational and molecular tools, provide a robust framework for systematically deciphering the complex logic of embryogenesis. This knowledge is invaluable for researchers and drug development professionals, as it opens new avenues for diagnosing and potentially treating a wide spectrum of congenital disorders.
A Researcher's Toolkit: Sequencing-Based and Imaging-Based Spatial Transcriptomic Platforms
Sequencing-based spatial transcriptomics (sST) represents a transformative set of technologies that enable the comprehensive mapping of gene expression within the context of intact biological tissues. Unlike traditional single-cell RNA sequencing that requires tissue dissociation, sST methods preserve the spatial coordinates of mRNA molecules, allowing researchers to correlate gene expression patterns with specific tissue architectures and cellular neighborhoods. This capability is particularly crucial for understanding complex biological systems where location dictates function, such as in embryonic development, tumor microenvironments, and organogenesis. The fundamental principle underlying all sST technologies involves assigning spatial barcodes to mRNA molecules directly in tissue sections, followed by high-throughput sequencing to simultaneously decode both gene identity and original location within the tissue [30] [31].
Two leading platforms in this field—10x Genomics' Visium and STOmics' Stereo-seq—exemplify different approaches to achieving spatial resolution while maintaining transcriptome-wide coverage. These platforms have enabled unprecedented insights into the spatial organization of gene expression and have become indispensable tools for researchers studying tissue architecture and cellular interactions. While both technologies ultimately generate spatial gene expression data, they differ significantly in their underlying biochemistry, resolution capabilities, and practical implementation. This technical guide examines the core methodologies of these platforms, with particular emphasis on their applications in embryonic research, where understanding the spatial distribution of mRNA is essential for unraveling the mechanisms of pattern formation and cell fate determination [32].
Core Technological Principles of Major sST Platforms
10x Genomics Visium Platform
The Visium platform employs a spatially barcoded oligonucleotide array patterned on glass slides to capture mRNA molecules from tissue sections. The fundamental workflow begins with tissue preservation and sectioning, followed by placement of tissue sections onto the Visium slides containing millions of capture spots with unique spatial barcodes. Each capture spot contains spatially barcoded oligo-dT primers that bind to polyadenylated mRNA molecules. After tissue permeabilization, which releases mRNA molecules from the tissue, the transcripts diffuse to the nearby capture spots and are bound by the barcoded primers. Reverse transcription then creates cDNA molecules that incorporate the spatial barcodes, effectively tagging each transcript with information about its original location in the tissue [30] [33].
The Visium platform has evolved through several iterations, with significant improvements in resolution and flexibility. The original Visium technology featured 55-μm barcoded spots with 100-μm center-to-center distances, providing resolution for approximately 1-10 cells per spot. The recently introduced Visium HD platform represents a substantial advancement, featuring a continuous lawn of 2-μm barcoded squares that enables single-cell-scale resolution. This enhanced architecture provides approximately 11 million unique spatial barcodes per capture area (6.5 × 6.5 mm), dramatically increasing spatial resolution while maintaining high sensitivity [30] [34]. The platform utilizes the CytAssist instrument to ensure high spatial fidelity by controlling reagent flow and minimizing lateral movement of transcripts during transfer from tissue to capture array, with studies demonstrating that 98.3-99% of transcripts remain localized within their expected morphological structures [34].
A key innovation in the Visium HD product line is the availability of two distinct chemistries tailored for different research applications. The Visium HD WT Panel uses probe-based chemistry optimized for differential gene expression analysis in human and mouse archival and FFPE samples, providing comprehensive coverage of protein-coding genes. In contrast, the Visium HD 3' chemistry employs a poly(A) capture-based approach that delivers whole transcriptome coverage, enabling expanded discovery applications including isoform detection, TCR/BCR analysis, and compatibility with diverse species beyond human and mouse [30] [33]. This flexibility makes the platform particularly valuable for embryonic research, where studying non-model organisms can provide crucial evolutionary insights.
STOmics Stereo-seq Platform
Stereo-seq (SpaTial Enhanced REsolution Omics-sequencing) employs a fundamentally different approach based on DNA nanoball (DNB) pattern arrays to achieve unprecedented spatial resolution. The technology utilizes a chip containing patterned arrays of DNBs, each approximately 200-500 nm in size, with each DNB containing unique spatial barcodes known as Coordinate IDs (CIDs). This nanoscale patterning enables Stereo-seq to achieve subcellular resolution (500 nm) while maintaining a large field of view that can be expanded up to decimeter scale (13 cm × 13 cm), a combination unique among current spatial transcriptomics technologies [31] [35].
The Stereo-seq workflow involves placing tissue sections onto the DNB chip array, where in situ RNA capture occurs through hybridization of polyadenylated RNA to the barcoded primers on the DNBs. After reverse transcription, the cDNA molecules are collected for library construction and sequencing. A critical advantage of this approach is that the spatial barcodes are incorporated during the array manufacturing process, allowing for extremely high barcode density and minimal distance between capture features. This high density enables the resolution of individual cells and even subcellular compartments, making it particularly powerful for studying intricate spatial patterns during embryonic development [31].
Another distinctive feature of Stereo-seq is its compatibility with multidimensional analysis, allowing integration of tissue staining with whole-genome mRNA transcriptomics and proteomics on the same tissue section. The platform's innovative staining approach enables researchers to correlate pathological features with spatiotemporal gene expression patterns, providing a more comprehensive understanding of tissue organization. Furthermore, Stereo-seq has demonstrated broad species compatibility, working effectively with cells, organs, and embryo sections from various animal and plant species, which is particularly valuable for evolutionary developmental biology studies comparing embryonic processes across species [31].
Figure 1: Comparative workflows of Visium and Stereo-seq platforms highlighting fundamental differences in array fabrication and tissue processing approaches.
Technical Comparison of Platform Capabilities
The selection between sST platforms requires careful consideration of technical specifications, experimental needs, and analytical requirements. The table below provides a comprehensive comparison of the key parameters for Visium, Visium HD, and Stereo-seq platforms:
| Parameter | Visium (v2) | Visium HD | Stereo-seq |
|---|---|---|---|
| Resolution | 55-μm spots (1-10 cells) | 2-μm squares (single-cell scale) | 500 nm (subcellular) |
| Spatial Barcode Density | ~5,000 features per area | ~11 million features per area | Nanoscale DNB patterning |
| Chemistry | Probe-based (WT) or poly(A) capture (3') | Probe-based (HD WT) or poly(A) capture (HD 3') | Poly(A) capture-based |
| Compatible Samples | FFPE, fresh frozen, fixed frozen | FFPE, fresh frozen, fixed frozen | Fresh frozen |
| Species Compatibility | Human, mouse (WT panel); Agnostic (3') | Human, mouse (HD WT); Broad species (HD 3') | Broad species compatibility |
| Field of View | 6.5 × 6.5 mm capture area | 6.5 × 6.5 mm capture area | Up to 13 × 13 cm |
| Spatial Fidelity | High (with CytAssist) | 98.3-99% transcript localization | Subcellular precision |
| Same-section Analysis | Gene expression, protein (IF), morphology (H&E) | Gene expression, protein (IF), morphology (H&E) | Gene expression, histology, proteomics |
| Sequencing Platform | Illumina-compatible | Illumina-compatible | DNBSEQ |
Table 1: Technical comparison of major sequencing-based spatial transcriptomics platforms, highlighting key differences in resolution, compatibility, and applications. Data compiled from [30] [31] [33].
The choice between these platforms involves important trade-offs. Visium HD offers single-cell-scale resolution while maintaining compatibility with FFPE samples, which is crucial for leveraging archival tissue banks in research. Its use of the CytAssist instrument provides a histology-friendly workflow that integrates with standard laboratory practices. In contrast, Stereo-seq provides superior subcellular resolution and an unprecedented field of view, enabling studies of large tissue areas or entire embryos at nanometer resolution. However, Stereo-seq is currently limited to fresh frozen samples, which may restrict its application to retrospectively collected specimens [30] [31] [35].
For embryonic research specifically, both platforms offer distinct advantages. The high resolution of Stereo-seq makes it ideal for mapping fine-grained expression patterns during early embryogenesis, such as the subcellular localization of maternal mRNAs that establish embryonic axes. Meanwhile, the flexibility of Visium HD in handling different sample types and its robust bioinformatics support may be advantageous for longer developmental time courses or when working with limited archival embryonic material. Recent applications in zebrafish and xenopus embryogenesis have demonstrated the power of these technologies for identifying spatially restricted genes and temporal expression dynamics during critical developmental windows [32].
Applications in Embryonic mRNA Localization Research
Sequencing-based spatial transcriptomics has revolutionized the study of embryonic development by enabling comprehensive mapping of mRNA localization patterns that establish the body plan. A prime example comes from research on early zebrafish embryogenesis, where enhanced tomo-seq (a related sST method) was used to systematically identify mRNA localization patterns in one-cell stage embryos. By embedding and orienting individual embryos along the animal-vegetal axis and cryosectioning them into 96 sections, researchers created high-resolution spatial transcriptomes that revealed three major groups of localized mRNAs: animal-pole localized, uniformly distributed, and vegetal-pole localized transcripts [32].
This approach led to the identification of 97 vegetally localized genes—a tenfold increase over previously known genes in this category—many of which play crucial roles in germ cell development and dorsoventral axis specification. The spatial transcriptomic data enabled researchers to follow the fate of these maternal transcripts through subsequent developmental stages using single-cell RNA metabolic labeling (scSLAM-seq), which distinguishes maternal and zygotic transcripts through 4-thiouridine incorporation. This combined approach revealed that many vegetally localized transcripts are specifically transported to primordial germ cells, demonstrating how sST technologies can uncover novel mechanisms of pattern formation in early embryos [32].
The power of sST technologies for embryonic research extends beyond zebrafish to other model organisms. Comparative analysis of mRNA localization in Xenopus laevis and Xenopus tropicalis oocytes using spatial transcriptomics has provided insights into the evolutionary conservation of localization mechanisms and identified enriched sequence motifs in localized transcripts. These studies highlight how sST enables systematic investigation of RNA transport and localization across vertebrate species, addressing fundamental questions about how initially pluripotent cells acquire spatial identity during embryogenesis [32].
For researchers studying embryonic development, sST platforms provide several unique capabilities. They enable transcriptome-wide analysis of spatial patterning without requiring prior knowledge of candidate genes, making them ideal for discovery-based approaches. They can resolve subtle expression gradients that establish embryonic axes and tissue boundaries. Furthermore, when applied across multiple timepoints, they can reconstruct developmental trajectories in both space and time, providing a four-dimensional understanding of embryogenesis [32].
Essential Research Reagents and Experimental Protocols
Key Research Reagent Solutions
Successful implementation of sST experiments requires careful selection of reagents and materials optimized for each platform. The following table outlines essential research reagent solutions and their functions in typical sST workflows:
| Reagent/Material | Function | Platform Specificity |
|---|---|---|
| Visium Spatial Gene Expression Slide | Contains spatially barcoded oligonucleotides for mRNA capture | 10x Genomics Visium specific |
| Visium HD Slide | Continuous lawn of 2-μm barcoded squares for enhanced resolution | 10x Genomics Visium HD specific |
| STOmics Stereo-seq Chip | DNA nanoball array with Coordinate IDs for spatial barcoding | STOmics Stereo-seq specific |
| CytAssist Instrument | Enables histology-friendly workflow and high spatial fidelity | 10x Genomics Visium (with FFPE samples) |
| Fixation and Permeabilization Reagents | Preserve tissue architecture while allowing mRNA diffusion to capture surface | Platform-independent (optimization required) |
| Reverse Transcription Mix | Converts captured mRNA to cDNA while incorporating spatial barcodes | Platform-specific formulations recommended |
| Library Preparation Kit | Prepares sequencing libraries from spatially barcoded cDNA | Platform-specific (Visium vs. Stereo-seq) |
| Histology Stains (H&E) | Visualize tissue morphology and guide region of interest selection | Platform-independent |
| 4-thiouridine (4sU) | Metabolic labeling for distinguishing maternal and zygotic transcripts | For temporal analyses in developing systems |
Table 2: Essential research reagent solutions for sequencing-based spatial transcriptomics experiments, highlighting platform-specific requirements.
Detailed Methodological Protocol for Embryonic sST
Based on published applications of sST in embryonic research, below is a detailed protocol for mapping mRNA spatial expression in early embryos:
Sample Preparation and Tissue Processing:
- Embryo Collection and Fixation: Collect embryos at desired developmental stages and immediately stabilize RNA while preserving spatial information. For zebrafish embryos, dechorionate if necessary and fix in 4% PFA for 24 hours at 4°C.
- Cryopreservation and Sectioning: Embed fixed embryos in OCT compound and orient along desired axes (e.g., animal-vegetal) using microscopic guidance. Section embryos at 10-20 μm thickness using a cryostat, ensuring complete sections for comprehensive spatial analysis.
- Tissue Placement and Permeabilization: Mount sections onto sST capture areas (Visium slides or Stereo-seq chips). Optimize permeabilization time to balance mRNA release efficiency with spatial resolution—typically 12-24 minutes using recommended permeabilization enzymes.
Spatial Library Preparation:
- mRNA Capture and Reverse Transcription: Perform on-slide reverse transcription using platform-specific reagents to convert captured mRNA to cDNA with incorporated spatial barcodes. For Visium, this occurs directly on the slide; for Stereo-seq, in situ capture is followed by cDNA collection.
- Second Strand Synthesis and Amplification: Generate double-stranded cDNA and amplify using PCR with platform-specific cycles to maintain representation while minimizing amplification bias.
- Library Construction and Sequencing: Fragment amplified cDNA, add platform-compatible adapters, and perform quality control before sequencing. Recommended sequencing depth is typically 50,000-100,000 reads per capture spot for Visium, adjusted accordingly for higher-resolution platforms.
Data Analysis and Integration:
- Spatial Expression Mapping: Use platform-specific computational tools (Space Ranger for Visium, STUtility for Stereo-seq) to align sequencing reads, count UMIs, and generate spatial expression matrices.
- Spatial Pattern Identification: Apply clustering algorithms and spatial autocorrelation analysis to identify spatially variable genes and expression patterns. For embryonic studies, focus on known patterning genes and novel spatially restricted transcripts.
- Validation and Integration: Confirm key findings using in situ hybridization or immunohistochemistry. Integrate with single-cell RNA-seq data when available to enhance cell type resolution [32] [36].
Figure 2: Experimental workflow for applying sequencing-based spatial transcriptomics to embryonic research, highlighting critical steps from embryo collection to developmental interpretation.
Data Analysis and Visualization Approaches
The analysis of sST data requires specialized computational approaches that simultaneously consider gene expression patterns and spatial relationships. The Seurat package has emerged as a widely used tool for analyzing spatially resolved transcriptomics data, extending its single-cell RNA-seq capabilities to incorporate spatial information. The typical analytical workflow begins with data normalization to account for technical variations while preserving biological signals. Unlike conventional scRNA-seq normalization methods that may assume uniform cell sizes, sST data requires specialized approaches like sctransform that effectively handle the substantial molecular count variance across spatial locations, which often reflects genuine biological heterogeneity rather than technical artifacts [36].
Following normalization, dimensionality reduction and clustering reveal spatially coherent cell states and tissue domains. However, unlike single-cell analysis where clusters represent cell types, sST clusters may correspond to spatially restricted domains or microenvironments. Seurat's spatial functions enable simultaneous visualization of clusters in both reduced dimension space (UMAP) and their physical tissue locations, facilitating the identification of anatomically meaningful patterns. The platform also provides tools for identifying spatially variable features—genes whose expression correlates with spatial location—using either pre-annotated anatomical regions or computational approaches like Moran's I that detect spatial autocorrelation without prior knowledge [36].
Effective visualization is crucial for interpreting sST data, particularly given the inherently spatial nature of the measurements. Basic approaches include SpatialFeaturePlot and SpatialDimPlot which overlay molecular data on tissue histology. More advanced visualization strategies address challenges such as color assignment for multiple cell types in complex tissues. The Spaco tool introduces a spatially aware colorization method that uses a "Degree of Interlacement" metric to model spatial relationships between categories, optimizing color assignments to enhance perceptual discrimination of neighboring cell types in intricate environments like embryonic tissues or brain regions [37]. Additional specialized visualizations include cell neighborhood analysis to characterize local interactions, transcript overlay maps to visualize individual RNA molecules, and multi-panel galleries that systematically highlight the spatial distributions of individual cell types or gene markers [38].
For embryonic research specifically, analytical workflows often incorporate temporal integration to reconstruct developmental trajectories across multiple timepoints. This enables researchers to connect early spatial expression patterns with later cell fate decisions, revealing how the spatial organization of gene expression guides morphogenesis. When combined with metabolic labeling approaches like scSLAM-seq, sST data can distinguish maternal and zygotic transcripts, enabling researchers to trace the fates of localized maternal mRNAs through critical developmental transitions such as zygotic genome activation and gastrulation [32].
Sequencing-based spatial transcriptomics has fundamentally transformed our ability to map gene expression within native tissue contexts, with particularly profound implications for developmental biology. As these technologies continue to evolve, several trends are likely to shape their future applications in embryonic research. First, the ongoing improvement in spatial resolution will enable increasingly precise mapping of gene expression patterns, potentially reaching the level of individual RNA molecules within subcellular compartments. Second, the integration of multi-omic measurements—combining spatial transcriptomics with proteomics, epigenomics, and metabolomics—will provide more comprehensive views of the molecular mechanisms governing pattern formation [31] [34].
The application of sST technologies to embryonic development has already yielded significant insights, from the systematic identification of localized maternal mRNAs in zebrafish to the evolutionary comparison of expression patterns across species. These studies demonstrate how sST moves beyond merely cataloging which genes are expressed to reveal how their spatial distribution creates functional organization within developing embryos. As these technologies become more accessible and analytical methods more sophisticated, they will undoubtedly uncover new principles of developmental biology and provide unprecedented views of the molecular architecture of embryogenesis [32].
For researchers embarking on sST studies of embryonic systems, the choice between platforms involves careful consideration of resolution requirements, sample compatibility, and analytical needs. Visium HD offers a robust, well-supported solution with single-cell resolution and FFPE compatibility, while Stereo-seq provides superior resolution and larger fields of view for comprehensive embryonic mapping. Whichever platform is selected, sST technologies promise to continue revolutionizing our understanding of how spatial patterns of gene expression guide the remarkable transformation from a single fertilized egg to a complex, multicellular embryo [30] [31] [32].
Imaging-based spatial transcriptomics has emerged as a revolutionary set of technologies that enable researchers to map gene expression patterns within their native spatial context in tissues and embryos. These methods overcome the fundamental limitation of traditional sequencing approaches, which require tissue dissociation and consequently lose all spatial information. For embryonic research, where the precise spatial organization of cells underlies fate determination and morphogenesis, these techniques provide an indispensable window into developmental processes. This technical guide details the core principles, methodologies, and applications of three prominent imaging-based spatial transcriptomics technologies: Multiplexed Error-Robust Fluorescence In Situ Hybridization (MERFISH), sequential Fluorescence In Situ Hybridization (seqFISH), and In Situ Sequencing (ISS). We provide comprehensive experimental protocols, quantitative performance comparisons, and practical implementation guidelines to equip researchers with the knowledge needed to apply these transformative techniques to the study of mRNA spatial expression in embryonic development.
The spatial organization of mRNA molecules within tissues and embryos serves as a critical regulator of cellular identity, function, and intercellular communication. During embryogenesis, the precise spatial and temporal control of gene expression patterns directs cell fate decisions, tissue patterning, and organ formation [39]. Traditional bulk and single-cell RNA sequencing methods, while powerful for cataloging transcriptional diversity, fundamentally destroy the spatial context of cells by requiring tissue dissociation [40] [39]. This limitation has driven the development of spatial transcriptomics technologies, which retain and record the spatial coordinates of RNA molecules within intact tissue samples.
Imaging-based spatial transcriptomics methods represent a particularly powerful subclass of these technologies, offering subcellular resolution and the ability to detect low-abundance transcripts that are often functionally significant in developmental processes [41] [42]. These techniques can be broadly categorized into two approaches: (1) in situ hybridization-based methods that use complementary nucleic acid probes to visualize RNA molecules, and (2) in situ sequencing-based methods that sequence RNA molecules directly within tissue sections [42]. This guide focuses on three prominent examples: MERFISH and seqFISH (representing in situ hybridization), and ISS (representing in situ sequencing).
The application of these techniques to embryonic research is particularly valuable. Embryos exhibit complex spatial patterns of gene expression that change dynamically over time, creating intricate molecular landscapes that guide morphogenesis. By preserving spatial context, these methods enable researchers to reconstruct gene expression gradients, identify signaling centers, trace cell lineages in situ, and uncover novel spatial relationships between cell types during critical developmental windows [43].
Technology-Specific Principles and Methodologies
MERFISH (Multiplexed Error-Robust Fluorescence In Situ Hybridization)
MERFISH is a massively multiplexed single-molecule imaging technology that combines single-molecule RNA FISH with combinatorial labeling and error-robust barcoding to simultaneously measure the copy number and spatial distribution of hundreds to thousands of RNA species in individual cells [40] [44]. The fundamental innovation of MERFISH is the assignment of a unique binary barcode to each RNA species, which is read out through sequential rounds of hybridization and imaging [40] [44].
The MERFISH workflow begins with probe design and hybridization. Encoding probes are designed to target specific RNA molecules, with each probe containing targeting sequences complementary to the RNA of interest and readout sequences that correspond to the bits of the RNA's binary barcode [45]. These encoding probes are hybridized to a fixed tissue sample, effectively imprinting the desired barcodes onto each RNA species.
Next, sequential rounds of hybridization and imaging are performed. In each round, fluorescently labeled readout probes are hybridized to their complementary readout sequences, imaged, and then removed through chemical cleavage [40] [45]. This process is repeated for multiple rounds, with each round corresponding to one bit in the binary barcode. The presence of a fluorescent spot indicates a "1" for that bit position, while its absence indicates a "0" [40].
The image processing and decoding phase follows, where computational algorithms identify fluorescent spots across all imaging rounds and decode their temporal barcodes into specific RNA identities [40]. Error-robust encoding schemes ensure accurate identification even in the presence of detection errors. Finally, cell segmentation is performed using nuclear (DAPI) and membrane staining to assign decoded RNAs to individual cells, generating spatial single-cell expression matrices [40].
MERFISH has demonstrated strong correlation with both bulk and single-cell RNA sequencing results, with improvements in overall dropout rates and sensitivity [40]. Recent advancements in MERFISH 2.0 have enhanced detection efficiency through optimized RNA anchoring, improved probe binding, and amplified readout probes, enabling nearly 100% detection efficiency with branched DNA amplification [44] [45].
seqFISH (sequential Fluorescence In Situ Hybridization)
seqFISH is a highly multiplexed spatial transcriptomics technology that identifies RNA transcripts through a fluorescent barcode built up over multiple rounds of hybridization and imaging [41] [46]. Unlike MERFISH's binary approach, seqFISH typically uses a larger palette of fluorescent colors in each round, enabling massive multiplexing with fewer rounds.
The core principle of seqFISH involves combinatorial labeling with multiple fluorophores. Each RNA species is assigned a unique barcode represented by a sequence of colors across multiple hybridization rounds [41] [46]. For example, a barcode might be "Round1-Red, Round2-Green, Round3-Blue" for one gene and "Round1-Green, Round2-Red, Round3-Red" for another.
The experimental workflow involves sequential hybridization cycles. In each cycle, fluorescently tagged readout probes are hybridized to their targets, imaged using multichannel fluorescence microscopy, and then chemically stripped [46]. This process is repeated for multiple cycles to build up the complete barcode for each transcript. The number of genes that can be distinguished grows exponentially with the number of rounds and colors per round.
A key advantage of seqFISH is its ability to achieve whole-transcriptome coverage. By using 60 rounds of hybridization with 4 colors each, researchers have demonstrated transcriptome-scale imaging of over 10,000 genes in single cells within tissues [47]. seqFISH also offers multi-omics capability, as it can be adapted to study nuclear architecture of DNA and combined with sequential immunofluorescence for protein detection [46].
seqFISH provides absolute quantification of RNA molecules, with each transcript appearing as a single dot in cells, and abundance quantified by counting these dots [41]. This approach avoids amplification biases inherent in sequencing-based methods and enables accurate detection of low-copy-number genes, including transcription factors and master regulators that are crucial in embryonic development [41].
ISS (In Situ Sequencing)
ISS represents a different approach to spatial transcriptomics, based on directly sequencing RNA molecules within tissue sections rather than decoding predetermined barcodes [42]. First published in 2013 by Nilsson's lab, ISS enables targeted readout of gene expression in fresh frozen tissues with spatial context [42].
The ISS workflow begins with tissue fixation and reverse transcription. mRNA in fixed tissues is reverse transcribed into complementary DNA (cDNA), preserving its spatial position [42]. Next, padlock probe hybridization and circularization occurs. Padlock probes are designed to target specific cDNA sequences; upon hybridization, they undergo circularization through ligation, forming circular DNA molecules [42].
Rolling circle amplification (RCA) follows, where the circularized padlock probes are amplified to create DNA nanoballs that remain co-localized with the original RNA molecules [42]. These amplified nanoballs serve as detectable landmarks for the original transcripts.
The barcoding and sequencing phase then occurs. ISS uses a sequencing-by-ligation (SBL) approach to decode the identity of each nanoball [42]. An anchor primer binds near the target sequence, and fluorescently labeled detection probes are ligated. Imaging after each ligation step determines the base identity at specific positions within the barcode. This process is repeated to build up the complete barcode sequence that identifies each mRNA species.
Variations of ISS have been developed to improve efficiency and applicability. HybISS (hybridization-based ISS) uses sequencing-by-hybridization to improve the signal-to-background ratio 5-fold [42]. SCRINSHOT avoids the reverse transcription step by employing SplintR ligase to directly ligate padlock probes to RNA, improving detection efficiency [42]. HybRISS uses chimeric padlock probes for direct RNA ligation, showing 5-fold improved efficiency compared to traditional ISS [42].
ISS has been used to detect up to 222 gene transcripts in various tissues and has proven particularly valuable in neuroscience applications, such as studying gene expression patterns in the vicinity of amyloid plaques in Alzheimer's disease models [42] [39].
Comparative Analysis of Technologies
Technical Performance Metrics
Table 1: Quantitative Comparison of Imaging-Based Spatial Transcriptomics Technologies
| Parameter | MERFISH | seqFISH | ISS |
|---|---|---|---|
| Multiplexing Capacity | Hundreds to tens of thousands of genes [44] | Up to 10,000+ genes demonstrated [47] | Hundreds of genes (up to 222 demonstrated) [42] |
| Detection Efficiency | Nearly 100% with bDNA amplification [45] | High efficiency with single-molecule sensitivity [41] | 5%-30% efficiency [42] |
| Spatial Resolution | Single-molecule, subcellular resolution [44] | Single-molecule, subcellular resolution [41] | Single-cell to subcellular resolution [42] |
| Tissue Compatibility | Diverse sample types including FFPE and frozen [44] | Virtually any tissue type [41] | Fresh frozen tissues [42] |
| Key Advantages | Error-robust barcoding, high reproducibility (R=0.99 between replicates) [40] | Whole-transcriptome coverage, multi-omics capability [46] | Robustness, commercial availability (10X Genomics) [42] |
| Primary Limitations | Requires predetermined gene panel | Multiple rounds of hybridization required | Moderate multiplexing capacity, optical crowding |
Workflow Diagrams
Figure 1: MERFISH Workflow. The method involves probe hybridization followed by multiple rounds of imaging to read out binary barcodes assigned to each RNA species.
Figure 2: seqFISH Workflow. The method uses multiple colors in each hybridization round to build fluorescent barcodes for transcript identification.
Figure 3: ISS Workflow. The method uses in situ sequencing via padlock probes and rolling circle amplification to identify transcripts.
Experimental Protocols for Embryonic Research
Sample Preparation for Embryonic Tissues
Proper sample preparation is critical for successful spatial transcriptomics in embryonic tissues, which often present unique challenges due to their small size, fragility, and dynamic nature. The following protocol is adapted for embryonic samples:
Tissue Collection and Fixation: Rapidly dissect embryos in cold PBS and immediately transfer to fresh, ice-cold 4% paraformaldehyde in PBS. Fix for 12-24 hours at 4°C with gentle agitation. The fixation time should be optimized based on embryo size and stage - shorter times (12-16 hours) for early embryos, longer times (18-24 hours) for later stages with more developed tissues.
Cryoprotection and Sectioning: Wash fixed embryos in PBS, then incubate in 30% sucrose solution at 4°C until embryos sink (typically 12-48 hours). Embed in OCT compound and rapidly freeze in dry ice-cooled isopentane. Section at 5-20μm thickness using a cryostat, collecting sections on poly-L-lysine coated coverslips suitable for imaging.
Permeabilization and Pre-hybridization: Treat sections with proteinase K (1-10μg/mL for 5-30 minutes at 37°C) based on embryo stage and size, followed by post-fixation in 4% PFA for 10 minutes. Permeabilize with 0.1-1% Triton X-100 in PBS for 30 minutes, then pre-hybridize in appropriate buffer to reduce background.
For MERFISH and seqFISH, RNA integrity is paramount. Assess RNA quality using RNA Integrity Number (RIN) with a minimum threshold of 4-6 for successful analysis [40]. For embryonic tissues with high RNase content (such as pancreas), additional precautions including RNase inhibitors in all solutions may be necessary [44].
MERFISH Protocol for Embryonic Tissues
Probe Hybridization: Apply encoding probes targeting your gene panel (typically 100-1000 genes for embryonic studies) in hybridization buffer. Use 100-200 encoding probes per RNA species to maximize signal [45]. Hybridize for 36-48 hours at 37°C in a humidified chamber.
Sequential Imaging Rounds:
- Perform 6-16 rounds of hybridization and imaging depending on the barcoding scheme
- For each round: hybridize fluorescent readout probes (30-60 minutes), image using a high-resolution fluorescence microscope with 20x-100x objective, then cleave fluorophores using 100mM DTT or TCEP
- Include DAPI and membrane stain (e.g., wheat germ agglutinin) in one round for cell segmentation
Image Processing and Analysis:
Recent improvements in MERFISH 2.0 chemistry, including enhanced RNA anchoring and amplified readout probes, can significantly improve performance in challenging embryonic tissues [44].
Data Analysis and Integration with Embryonic Atlases
The analysis of spatial transcriptomics data from embryonic samples requires specialized computational approaches:
Spatial Expression Pattern Identification: Use spatial autocorrelation statistics (Moran's I, Geary's C) to identify genes with non-random spatial distributions. Cluster cells based on spatial expression patterns to identify tissue domains and emerging structures.
Developmental Trajectory Inference: Integrate with temporal data to reconstruct spatial developmental trajectories using methods such as RNA velocity or pseudotime analysis adapted to spatial contexts.
Cell-Cell Communication Analysis: Infer signaling interactions between spatially proximal cells using tools like NicheNet or CellPhoneDB, adapted to account for spatial constraints.
Registration with Reference Atlases: Align spatial expression data with anatomical reference atlases of embryonic development to place molecular profiles within established morphological contexts.
Research Reagent Solutions
Table 2: Essential Research Reagents for Imaging-Based Spatial Transcriptomics
| Reagent Category | Specific Examples | Function | Technology Application |
|---|---|---|---|
| Probes | Encoding probes with readout sequences [45] | Target-specific RNA detection through barcode assignment | MERFISH |
| Readout probes with cleavable fluorophores [45] | Fluorescent barcode readout in sequential rounds | MERFISH, seqFISH | |
| Padlock probes [42] | Target recognition and circularization for RCA | ISS | |
| Amplification Components | Branched DNA (bDNA) amplifiers [45] | Signal amplification for improved detection efficiency | MERFISH |
| Rolling circle amplification (RCA) enzymes [42] | Signal amplification via DNA nanoball formation | ISS | |
| Stains and Labels | DAPI | Nuclear counterstain for cell segmentation | All technologies |
| Membrane stains (WGA, antibodies) [40] | Cell boundary delineation for segmentation | All technologies | |
| Specialized Buffers | Hybridization buffers | Optimal probe binding conditions | All technologies |
| Cleavage buffers (DTT, TCEP) [40] | Fluorophore removal between imaging rounds | MERFISH, seqFISH | |
| Commercial Platforms | Vizgen MERSCOPE Platform [44] | Integrated instrument for MERFISH | MERFISH |
| Spatial Genomics GenePS [46] | seqFISH platform | seqFISH | |
| 10X Genomics Visium [42] | Commercial ISS platform | ISS |
Imaging-based spatial transcriptomics technologies represent a powerful toolkit for investigating mRNA spatial expression in embryonic development. MERFISH, seqFISH, and ISS each offer unique advantages and considerations for developmental biologists. MERFISH provides high accuracy and reproducibility through error-robust barcoding, making it ideal for targeted studies of specific gene networks. seqFISH offers unparalleled multiplexing capacity, enabling whole-transcriptome mapping critical for discovery-based approaches in complex developing systems. ISS delivers robust targeted detection with commercial accessibility, suitable for focused investigations of known patterning genes.
For embryonic research specifically, the selection of an appropriate spatial transcriptomics method depends on several factors: the developmental stage, the number of genes of interest, required spatial resolution, and available resources. Early embryos with rapidly changing gene expression patterns may benefit from the whole-transcriptome coverage of seqFISH, while later stages with established tissue compartments might be effectively studied with targeted MERFISH panels focused on specific differentiation programs.
As these technologies continue to advance, we anticipate further improvements in detection efficiency, multiplexing capacity, and accessibility. Emerging directions include multi-omic integration (simultaneous RNA and protein detection), live-cell spatial transcriptomics to capture dynamics, and computational methods for reconstructing developmental trajectories from spatial data. By applying these sophisticated tools to embryonic systems, researchers can unlock new understanding of how spatial patterns of gene expression orchestrate the remarkable process of development from a single cell to a complex organism.
The process of embryogenesis represents one of biology's most complex and precisely orchestrated events, where initially pluripotent cells differentiate into diverse cell types with distinct spatial organization. During early embryonic development, particularly in model organisms like zebrafish and Xenopus, the subcellular localization and transport of maternal mRNA fundamentally guides cell fate decisions and axis formation [32]. However, this critically important spatial information is completely lost in conventional bulk RNA sequencing and single-cell RNA sequencing (scRNA-seq) due to tissue dissociation, which disrupts the native structural organization of the developing embryo [3].
Spatial transcriptomics has emerged as a revolutionary technological field that enables researchers to map gene expression within intact tissues, preserving the precise spatial context of mRNA molecules [48]. For embryonic research, this means researchers can now visualize the spatial distribution of RNA molecules and gain unprecedented insights into cellular organization, interactions, and functions in their native developmental environment [3]. The ability to precisely locate where genes are expressed within highly structured embryonic tissues is essential for elucidating fundamental biological processes including gene functions, gene-gene interactions, cell-cell communication, dynamic molecular processes, and microenvironmental influences that drive development [3].
A practical challenge facing embryology researchers today is the proliferation of diverse spatial transcriptomics technologies, each with distinct technical parameters including cellular resolution, detection sensitivity, gene coverage, and throughput [3]. This comprehensive guide provides an in-depth technical comparison of major spatial transcriptomics platforms, with specific application to embryonic research contexts, to empower researchers in selecting the optimal technology for their specific developmental biology questions.
Spatial transcriptomics technologies can be broadly categorized into two fundamental methodological groups: sequencing-based and imaging-based approaches [3] [49]. While both aim to reveal the spatial locations of gene expression, their underlying technologies differ dramatically in how they capture spatial information and determine abundance of specific mRNA molecules within tissue [3].
Sequencing-Based Spatial Transcriptomics
Sequencing-based technologies integrate spatially barcoded arrays with next-generation sequencing to determine transcript locations and expression levels within tissues [3]. These methods generally work by capturing mRNA transcripts from tissue sections using polyT tails built into unique, spatially barcoded probes on an array [3]. During cDNA synthesis, these spatial barcodes are incorporated into each cDNA molecule, and through subsequent library construction and sequencing, researchers can determine expression levels and map them back to precise locations based on their spatial barcodes [3].
The fundamental difference among sequencing-based technologies primarily lies in the feature size of the array, which largely determines spatial resolution [3]. Platforms in this category include 10X Visium, Visium HD, and Stereo-seq, with the GeoMx Digital Spatial Profiler employing a combination strategy of barcoded probes and region-of-interest selection [3].
Imaging-Based Spatial Transcriptomics
Imaging-based technologies employ single-molecule fluorescence in situ hybridization (smFISH) as their backbone technology [3]. These methods enable simultaneous detection of hundreds to thousands of RNA transcripts through cyclic, highly multiplexed smFISH [3]. This is achieved using primary probes that hybridize to specific RNA transcripts, followed by secondary probes labeled with different fluorophores that bind to the primary probes [3].
By sequentially hybridizing and imaging fluorescence from these secondary probes, researchers can determine spatial location and expression levels of individual RNA transcripts based on transcript-specific fluorescent signatures and their intensity [3]. The differences between imaging-based platforms are mainly in probe design, hybridization protocols, signal amplification methods, and gene decoding strategies [3]. Major platforms in this category include Xenium, Merscope, and CosMx [3].
Table 1: Fundamental Differences Between Sequencing and Imaging Approaches
| Parameter | Sequencing-Based | Imaging-Based |
|---|---|---|
| Core Technology | Spatially barcoded arrays + NGS [3] | Multiplexed FISH [3] |
| Spatial Resolution | Multi-cell to single-cell [49] | Single-cell to subcellular [49] |
| Gene Coverage | Whole transcriptome, unbiased [49] | Targeted panels (hundreds to thousands) [49] |
| Throughput | Higher slide throughput [49] | Lower slide throughput [49] |
| Data Output | Expression matrices with spatial coordinates [49] | Large image datasets requiring processing [49] |
| Best Application | Discovery-driven research [49] | Validation and high-resolution localization [49] |
Spatial Transcriptomics Technology Decision Tree
Technical Comparison of Major Commercial Platforms
Imaging-Based Platforms: Xenium, Merscope, and CosMx
Xenium employs a hybrid technology combining in situ sequencing (ISS) and in situ hybridization (ISH) [3]. The process involves padlock probes containing gene-specific barcodes hybridizing to target RNA transcripts, followed by ligation and rolling circle amplification (RCA) to enhance signal sensitivity [3]. Fluorescently labeled oligonucleotide probes then bind to gene-specific barcodes, with successive rounds of hybridization using different fluorophores generating unique optical signatures for each target gene [3].
Merscope utilizes a distinctive binary barcode strategy for gene identification [3]. Each gene is assigned a unique binary barcode consisting of a sequence of "0"s and "1"s [3]. Thirty to fifty gene-specific primary probes hybridize to different regions of the target gene, with fluorescently labeled secondary probes reading the barcode over multiple imaging rounds [3]. Fluorescence detection corresponds to "1" in the barcode, while absence indicates "0" [3]. This binary barcoding strategy reduces optical crowding and supports error correction [3].
CosMx employs a hybridization method similar to MERSCOPE but incorporates an additional positional dimension for gene identification [3]. The process begins with a pool of five gene-specific probes containing target-binding domains and readout domains with 16 sub-domains [3]. Each secondary probe includes a primary probe binding domain linked to a branched, fluorescently labeled readout domain through a UV-cleavable linker [3]. The combination of four fluorescent colors and 16 sub-domains generates unique color and position signatures for each target gene, enabling identification of more targets than Xenium and MERSCOPE [3].
Sequencing-Based Platforms: Visium, Visium HD, and Stereo-seq
10X Visium relies on spatially barcoded RNA-binding probes attached to slides, containing spatial barcodes for location decoding, unique molecular identifiers (UMIs) for transcript quantification, and oligo-dT sequences for mRNA binding [3]. Visium offers two workflow versions: V1 for fresh tissue and V2 for both fresh and FFPE tissue with a modified mRNA capture strategy requiring a CytAssist instrument [3].
Visium HD uses the same technology as Visium V2 workflow but features significantly smaller spot size of 2 μm compared to the standard 55 μm feature size in original Visium, dramatically enhancing spatial resolution [3].
Stereo-seq utilizes DNA nanoball (DNB) technology for in situ RNA capture [3]. Unlike 10X Visium, which directly attaches oligo probes to slides, Stereo-seq oligo probes are circularized and used as templates to generate DNA nanoballs via rolling circle amplification [3]. The DNBs are loaded onto a grid-patterned array with a diameter of approximately 0.2 μm and center-to-center distance of 0.5 μm, significantly smaller than the 2 μm spots in Visium HD [3].
Table 2: Comprehensive Platform Comparison for Embryonic Research Applications
| Platform | Technology Type | Spatial Resolution | Gene Coverage | Tissue Compatibility | Embryonic Research Applications |
|---|---|---|---|---|---|
| 10X Visium | Sequencing-based [3] | 55 μm spots (multi-cell) [3] | Whole transcriptome [49] | FF, FFPE [3] | Tissue-level patterning, domain identification |
| Visium HD | Sequencing-based [3] | 2 μm spots (single-cell) [3] | Whole transcriptome [3] | FF, FFPE [3] | Single-cell mapping in complex embryonic tissues |
| Stereo-seq | Sequencing-based [3] | 0.5 μm center-to-center (subcellular) [3] | Whole transcriptome [3] | FF [3] | High-resolution developmental atlas construction |
| Xenium | Imaging-based [3] | Subcellular [49] | Targeted panels (hundreds of genes) [3] | FF, FFPE [3] | Subcellular mRNA localization in early embryos |
| Merscope | Imaging-based [3] | Subcellular [3] | Targeted panels (hundreds to thousands) [3] | FF, FFPE [3] | Error-resistant transcript mapping in development |
| CosMx | Imaging-based [3] | Subcellular [48] | Targeted panels (whole transcriptome available) [48] | FF, FFPE [48] | High-plex spatial phenotyping in embryonic structures |
| GeoMx DSP | Sequencing-based [3] | 10-600 μm ROI selection [48] | Whole transcriptome [48] | FFPE [48] | Region-specific analysis in complex embryonic structures |
Experimental Design and Workflow Considerations for Embryonic Research
Workflow Diagrams for Spatial Transcriptomics Technologies
Sequencing vs Imaging Experimental Workflows
Special Considerations for Embryonic Tissues
Embryonic tissues present unique challenges for spatial transcriptomics analysis. They are often delicate, with heterogeneous cell densities and complex three-dimensional architecture that evolves rapidly through developmental time [32]. For early embryonic stages, particularly in zebrafish and Xenopus models, researchers have successfully adapted methods like tomo-seq to systematically investigate spatial RNA gradients [32].
In tomo-seq applications for embryonic research, individual embryos are embedded and oriented along visible axes (e.g., animal-vegetal axis), then sectioned into numerous thin sections (e.g., 96 sections) for processing [32]. This approach has enabled identification of distinct mRNA localization patterns, including transcripts specifically localized to the vegetal pole that play crucial roles in germ cell development and axis specification [32].
For imaging-based approaches, embryonic tissues may require optimization of permeabilization conditions and hybridization times due to differences in extracellular matrix composition compared to adult tissues. Sequencing-based methods must account for potentially lower RNA yields from small embryonic structures while maintaining spatial integrity.
Data Analysis and Computational Tools for Embryonic Spatial Transcriptomics
The computational analysis of spatial transcriptomics data requires specialized tools that can integrate spatial information with gene expression patterns. For embryonic research, this often involves mapping developmental trajectories and identifying spatially restricted gene expression programs.
Thor represents an advanced platform specifically designed for cell-level investigation of spatial transcriptomics that integrates histological image analysis [50]. This multi-modal approach is particularly valuable for embryonic tissues where cellular morphology provides important cues about developmental states. Thor employs an anti-shrinking Markov diffusion method to infer single-cell spatial transcriptome from spot-level data, effectively combining gene expression and cell morphology [50]. The platform includes modular tools for genomic and image-based analysis, enabling comprehensive tissue interpretation in spatial biology [50].
PERSIST (PredictivE and Robust gene SelectIon for Spatial Transcriptomics) provides a flexible deep learning framework to identify informative gene targets for spatial transcriptomics studies by leveraging reference scRNA-seq data [51]. This approach is particularly valuable for embryonic research where specific developmental processes may require customized gene panels. PERSIST reliably identifies panels that provide more accurate prediction of genome-wide expression profiles, capturing more information with fewer genes [51]. The method's binarization of gene expression levels enables models trained on scRNA-seq data to generalize to spatial transcriptomics data despite the complex technological shift [51].
Table 3: Essential Research Reagent Solutions for Embryonic Spatial Transcriptomics
| Reagent/Category | Function | Platform Compatibility | Embryonic Research Considerations |
|---|---|---|---|
| Spatially Barcoded Beads/Slides | Capture mRNA with spatial coordinates | 10X Visium, Visium HD, Stereo-seq [3] | Optimal orientation of embryonic axes critical |
| Multiplexed FISH Probes | Hybridize to target transcripts for imaging | Xenium, Merscope, CosMx [3] | Panel design should include developmental markers |
| Tissue Permeabilization Enzymes | Enable probe or reagent access to mRNA | All platforms | Concentration may need optimization for embryonic tissues |
| Nucleotide Analogs (4sU/4sUTP) | Metabolic labeling for temporal analysis | scSLAM-seq adaptations [32] | Enables tracking maternal vs. zygotic transcripts |
| Cryo-embedding Media | Tissue preservation for sectioning | All platforms | Orientation of small embryos requires precision |
| UV-Cleavable Indexes | Spatially restricted release of barcodes | GeoMx DSP [48] | ROI selection based on embryonic morphology |
| RNase Inhibitors | Preserve RNA integrity during processing | All platforms | Particularly critical for RNA-rich embryonic tissues |
Application to Embryonic mRNA Localization and Developmental Research
The application of spatial transcriptomics to embryonic development has already yielded significant insights into the spatial organization of gene expression during critical developmental processes. Research in zebrafish embryos has demonstrated how spatially resolved transcriptomics combined with single-cell RNA labeling can track the fate of individual maternal transcripts until gastrulation [32].
This approach has revealed that many transcripts localized to the vegetal pole are specifically transported to primordial germ cells, providing mechanistic insights into germ cell specification [32]. Furthermore, comparative spatial transcriptome analysis across species (zebrafish and Xenopus) has illuminated evolutionary conservation of localized genes and enriched sequence motifs [32].
For embryonic research, the choice between sequencing-based and imaging-based spatial transcriptomics often depends on the specific developmental question. Sequencing-based approaches are ideal for discovery-driven research where the goal is to identify novel spatially patterned genes without prior assumptions [49]. This makes them particularly valuable for constructing comprehensive spatial maps of embryonic gene expression at key developmental stages.
Imaging-based approaches excel when high spatial resolution is required to resolve subcellular localization patterns or when focusing on a predefined set of candidate genes with known relevance to development [49]. These methods are particularly powerful for analyzing early embryonic stages where precise subcellular localization of maternal mRNAs dictates developmental outcomes.
The rapidly evolving landscape of spatial transcriptomics technologies presents both opportunities and challenges for developmental biologists studying embryonic mRNA localization. Current platforms offer complementary strengths, with sequencing-based methods providing unbiased whole-transcriptome coverage and imaging-based methods delivering superior spatial resolution.
For embryonic research, the optimal platform choice depends on multiple factors including the developmental stage, species, specific biological question, and available resources. Sequencing-based approaches like Visium HD and Stereo-seq offer compelling options for building comprehensive spatial maps of embryonic gene expression, while imaging-based platforms like Xenium and Merscope provide the resolution needed to analyze subcellular mRNA localization patterns critical for early development.
Future directions in the field will likely include increased integration of spatial transcriptomics with live imaging approaches, enhanced computational methods for reconstructing three-dimensional expression patterns from two-dimensional sections, and development of specialized workflows optimized for the unique challenges of embryonic tissues. As spatial technologies continue to advance in resolution, sensitivity, and throughput, they will undoubtedly yield unprecedented insights into the spatial orchestration of gene expression that guides the remarkable process of embryonic development.
The process of organogenesis, where embryonic cells differentiate and organize into complex tissues and organs, is governed by precise spatiotemporal gene expression patterns. Traditional bulk and single-cell RNA sequencing (scRNA-seq) have been instrumental in identifying cellular heterogeneity but fundamentally lack the spatial context essential for understanding developmental biology [52]. The loss of this spatial information is a critical limitation, as the spatial localization of cells and molecules, along with intercellular interactions within the tissue context, are pivotal for elucidating how signaling networks orchestrate morphogenesis [52]. Spatial transcriptomics (ST) has revolutionized this landscape by simultaneously capturing gene expression profiles and in situ spatial information of tissues, thereby providing a visual spatial transcriptome map integrated with tissue morphological characteristics [52].
This technical guide explores the application of spatial transcriptomics in mapping organogenesis and the signaling networks that underpin it, with a specific focus on embryonic development. The content is framed within the broader thesis of detecting mRNA spatial expression in embryo research, providing researchers, scientists, and drug development professionals with a detailed overview of the technologies, methodologies, and analytical frameworks driving this field.
Core Spatial Transcriptomics Technologies and Selection Criteria
Spatial transcriptomics technologies can be broadly classified into two categories based on RNA detection strategies: imaging-based and sequencing-based methods [52] [27] [3]. The choice between them depends on the specific research questions, desired resolution, and required gene coverage.
Imaging-Based Spatial Transcriptomics
Imaging-based technologies utilize in situ hybridization (ISH) or in situ sequencing (ISS) to detect and localize RNA molecules directly within intact tissue sections.
- ISH-based Technologies (e.g., MERFISH, RNAscope): These methods use fluorescently labeled probes complementary to target RNA sequences. MERFISH employs a unique binary barcoding strategy with error correction, significantly enhancing the robustness of transcript recognition [52] [3]. RNAscope uses "double-Z" probes that must hybridize adjacently on the target RNA, providing high sensitivity and specificity for detecting low-abundance transcripts at subcellular resolution [27]. Commercial platforms like the CosMx SMI combine a MERFISH-like strategy with an optical signature approach, using multiple rounds of hybridization with fluorescent readouts to detect thousands of transcripts [27] [3].
- ISS-based Technologies (e.g., Xenium, STARmap): These techniques involve the direct sequencing of RNA in its native tissue context. Xenium, a commercial platform, is a hybrid technology that uses padlock probes and rolling circle amplification (RCA) to generate signals for targeted genes with high sensitivity and specificity [52] [3]. STARmap also uses barcoded padlock probes and RCA but embeds the products in a hydrogel, allowing for 3D spatial mapping in thick tissue samples [27].
A key advantage of imaging-based methods is their high spatial resolution, often at a subcellular level, and their excellent performance with formalin-fixed, paraffin-embedded (FFPE) samples, which is valuable for working with archival clinical specimens or hard tissues like bone [52]. A limitation is that most are targeted approaches, requiring a predefined set of genes for probe design, which can restrict the discovery of novel transcripts [52].
Sequencing-Based Spatial Transcriptomics
Sequencing-based technologies rely on capturing mRNA onto spatially barcoded arrays followed by next-generation sequencing (NGS).
- 10X Visium & Visium HD: The Visium platform uses a slide coated with spots containing spatially barcoded oligo-dT probes. During tissue permeabilization, released mRNA binds to these probes, and after cDNA synthesis and sequencing, expression data is mapped back to the spatial barcodes [3]. The original Visium has a spot resolution of 55 μm, while the newer Visium HD reduces this to a 2 μm feature size, dramatically enhancing resolution [3].
- Stereoseq: This platform uses DNA nanoball (DNB) technology. Oligo probes are circularized and amplified into DNBs, which are then patterned onto an array at a very high density, achieving a submicrometer center-to-center distance and thus providing extremely high spatial resolution [3].
These methods offer the advantage of unbiased, whole-transcriptome coverage, making them ideal for exploratory discovery research. However, their initial spatial resolution was lower than imaging-based methods, a gap that newer platforms like Visium HD and Stereoseq are actively closing [3].
Table 1: Comparison of Major Commercial Spatial Transcriptomics Platforms
| Technology | Core Method | Spatial Resolution | Transcript Coverage | Key Advantage | Key Limitation |
|---|---|---|---|---|---|
| 10X Visium HD [3] | Sequencing-based | 2 μm | Whole transcriptome | High resolution, unbiased discovery | Requires CytAssist instrument for FFPE |
| Xenium [52] [3] | Imaging-based (ISS/ISH) | Subcellular | Targeted (up to 5,000 genes) | High sensitivity & specificity, fast data output | Targeted approach limits novel discovery |
| MERFISH [52] [3] | Imaging-based (ISH) | Subcellular | Targeted (thousands of genes) | High sensitivity, binary barcode error-correction | Complex probe design and imaging |
| CosMx SMI [27] [3] | Imaging-based (ISH) | Subcellular | Targeted (up to 19,000 transcripts) | High-plex protein co-detection, high-plex RNA | High cost, complex workflow |
| Stereoseq [3] | Sequencing-based (DNB) | ~0.5 μm | Whole transcriptome | Ultra-high resolution, unbiased discovery | Less established, data complexity |
Experimental Protocol for Embryonic Spatial Transcriptomics
The following protocol outlines a generalized workflow for applying spatial transcriptomics to embryonic tissues, integrating steps from various successful studies [52] [53].
Sample Preparation and Fixation
- Tissue Collection: Isolate mouse embryos at the desired developmental stage (e.g., E7.5 for gastrulation studies [53]). Precise developmental staging is critical.
- Fixation: Immediately immerse the embryo in 4% paraformaldehyde (PFA) for 24 hours at 4°C. This cross-links biomolecules and preserves tissue architecture and RNA integrity.
- Dehydration and Embedding:
- For FFPE samples: Dehydrate the fixed tissue through a graded ethanol series, clear with xylene, and embed in paraffin wax. Section at 5-10 μm thickness.
- For fresh frozen samples: Cryoprotect the fixed tissue by incubating in 30% sucrose solution until it sinks. Embed in Optimal Cutting Temperature (O.C.T.) compound and rapidly freeze on dry ice. Section at 10-20 μm thickness using a cryostat.
On-Slide Processing for Sequencing-Based ST (e.g., 10X Visium)
- Deparaffinization and Staining: For FFPE sections, deparaffinize with xylene and rehydrate through a graded ethanol series. Stain with Hematoxylin and Eosin (H&E) and image the slide using a brightfield microscope for histological reference.
- mRNA Capture: Permeabilize the tissue to allow mRNA to diffuse and bind to the spatially barcoded oligos on the Visium slide. The V2 workflow for FFPE samples uses gene-specific probes that are hybridized to the RNA and then captured via ligation onto the slide's poly(dT) probes [3].
- cDNA Synthesis and Library Construction: Synthesize cDNA on the slide. Harvest the cDNA, which now contains the spatial barcodes and unique molecular identifiers (UMIs). Proceed with standard NGS library preparation, including amplification and index addition.
On-Slide Processing for Imaging-Based ST (e.g., Xenium)
- Probe Hybridization: Apply a panel of gene-specific padlock probes to the tissue section. These probes hybridize to their target mRNA sequences.
- Ligation and Amplification: If both ends of a padlock probe hybridize adjacently to the target, they are ligated to form a circular DNA molecule. This circle is then amplified via RCA, creating a rolling circle product (RCP) that contains multiple copies of the gene-specific barcode [3].
- Fluorescent Readout and Imaging: Add fluorescently labeled readout probes that bind to the barcodes within the RCPs. Image the slide using a high-resolution fluorescence microscope. The fluorophores are then cleaved off, and the process is repeated for multiple rounds to decode the optical signatures of each target gene [52] [3].
Data Analysis and Integration
- Alignment and Gene Expression Matrix: For sequencing-based data, demultiplex sequencing reads and align them to a reference genome to create a gene expression matrix where each row is a gene and each column is a spatial barcode.
- Cell Segmentation: Use the companion H&E image (for sequencing-based) or DAPI staining (for imaging-based) to identify individual cell boundaries. This allows for assigning transcripts to specific cells.
- Clustering and Annotation: Perform dimensionality reduction and clustering on the spatially resolved gene expression data. Annotate cell types based on known marker genes. In a study of mouse gastrulation, this process resolved over 80 refined cell types across germ layers [53].
- Integration with scRNA-seq Data: Use computational tools to integrate ST data with a pre-existing scRNA-seq reference atlas. This enhances cell type annotation and can help infer the location of cell types that are less abundant in the ST data [53].
- Spatial Analysis: Identify spatially variable genes, reconstruct cell-cell communication networks based on ligand-receptor co-expression in neighboring cells, and map the expression gradients of key morphogens.
Diagram Title: Spatial Transcriptomics Workflow for Embryonic Tissues
Mapping Signaling Networks in Embryonic Development
Spatial transcriptomics provides an unprecedented view of how signaling pathways pattern the embryo. A key application is the construction of a spatiotemporal atlas. For example, an integrated atlas of mouse gastrulation from E6.5 to E9.5 combined ST with scRNA-seq data from over 150,000 cells, enabling the exploration of gene expression dynamics across the anterior-posterior and dorsal-ventral axes and uncovering the spatial logic guiding mesodermal fate decisions in the primitive streak [53].
The Nodal and BMP Pathways in Patterning
The TGF-β family morphogens, including Nodal and BMP, are critical for germ layer specification and dorsal-ventral (D/V) patterning during gastrulation [54]. Studies in zebrafish have revealed nuanced roles for the central effector Smad4. Contrary to the classical view, loss of Smad4 disrupts BMP signaling and D/V patterning entirely but maintains Nodal signaling, albeit with reduced robustness. This Smad4-independent Nodal signaling is sufficient for mesoderm specification but not for optimal endoderm specification, demonstrating how signaling robustness and specificity are differentially controlled within the same network [54].
Signaling in Pre-implantation Embryos
In early mammalian development, signaling pathways precisely orchestrate the first lineage segregations. The Hippo pathway is a key regulator of the fate decision between the inner cell mass (ICM) and the trophectoderm (TE). In outer, polarized cells of the morula, the Hippo pathway is inactive, allowing the transcriptional coactivators YAP/TAZ to enter the nucleus and, with TEAD4, activate TE-specific genes like CDX2. In inner, non-polar cells, the active Hippo pathway leads to phosphorylation and cytoplasmic retention of YAP/TAZ, promoting ICM fate [55]. Other pathways like Wnt/β-catenin, FGF, and TGF-β/Activin/Nodal are also involved in fine-tuning the specification of the epiblast (EPI) and primitive endoderm (PrE) within the ICM [55].
Diagram Title: Hippo Pathway in Early Lineage Specification
Table 2: Experimental Modulation of Key Signaling Pathways in Pre-implantation Embryos [55]
| Small Molecule / Ligand | Target Pathway | Effect (A./I.) | Key Phenotypic Outcome on Blastocyst |
|---|---|---|---|
| TRULI | Hippo | Inhibition (I.) | Increased ICM markers, decreased TE markers |
| FGF2 | FGF | Activation (A.) | Decreased EPI, increased PrE markers |
| PD173074 | FGF | Inhibition (I.) | Increased EPI, decreased PrE markers |
| SB431542 | TGF-β/Nodal | Inhibition (I.) | Increased EPI markers, no change in PrE |
| Activin A | TGF-β/Nodal | Activation (A.) | No significant change in EPI or PrE markers |
| BMP4 | BMP | Activation (A.) | Severely reduced blastocyst development rate |
Table 3: Key Research Reagent Solutions for Spatial Transcriptomics
| Item | Function / Application | Example Use Case |
|---|---|---|
| Padlock Probes [3] | Target-specific probes for ISS methods (e.g., Xenium). Hybridize to mRNA and are circularized for RCA. | Detecting a predefined panel of genes involved in a specific signaling pathway (e.g., Nodal targets). |
| Spatially Barcoded Beads/Slides [52] [3] | Capture mRNA with spatial context for sequencing-based ST (e.g., 10X Visium). | Unbiased, whole-transcriptome mapping of an embryonic tissue section. |
| Fluorescently Labeled Readout Probes [3] | Bind to barcodes in amplified products for cyclic imaging and signal decoding. | Visualizing the spatial distribution of hundreds to thousands of RNA species in a single sample. |
| CytAssist Instrument [3] | (10X Visium) Transfers probes from a standard slide to the Visium slide, optimizing workflow for FFPE samples. | Enabling spatial genomics on archived clinical embryo specimens or hard-to-process tissues. |
| Nucleic Acid Dyes (DAPI) | Counterstain for cell segmentation and nuclear localization in imaging-based ST. | Defining cell boundaries to assign transcripts to individual cells during data analysis. |
| Specific Pathway Modulators [55] | Small molecules to activate or inhibit key pathways (see Table 2). | Functionally testing the role of a pathway (e.g., Hippo, FGF) in lineage specification. |
Spatial transcriptomics has fundamentally transformed our ability to decode the complex processes of organogenesis and the signaling networks that govern them. By moving beyond mere cataloging of cell types to revealing their precise spatial organization and communication, ST provides a dynamic map from which developmental mechanisms can be directly inferred and experimentally tested. The integration of ST with single-cell atlases, as demonstrated in the spatiotemporal atlas of mouse gastrulation, offers a powerful framework for projecting in vitro models, such as gastruloids, onto an in vivo reference space [53]. For researchers and drug developers, these technologies and methodologies provide the tools to not only understand the fundamental logic of life's building process but also to identify the spatial etiologies of developmental disorders and pioneer novel therapeutic strategies. As technologies advance towards higher resolution, 3D reconstruction, and multi-omic integration, the journey from data to discovery in embryology is poised to accelerate dramatically.
Navigating Technical Challenges: From Sample Preparation to Data Fidelity
Optimizing Tissue Preservation, Sectioning, and Permeabilization for Embryonic Tissues
The precise mapping of mRNA spatial expression in embryonic tissues is fundamental to understanding developmental biology, cellular differentiation, and morphogenetic events. This technical guide outlines optimized protocols for tissue preservation, sectioning, and permeabilization specifically contextualized within a broader thesis on detecting mRNA spatial expression in embryo research. The integrity of spatial gene expression data is profoundly influenced by pre-analytical variables including fixation chemistry, sectioning parameters, and permeabilization efficiency. For embryonic tissues, which often exhibit heightened fragility and dynamic transcriptional activity, these procedures require specialized modifications to preserve both morphological detail and RNA integrity. The methods described herein are designed to support high-resolution spatial transcriptomic analyses, enabling researchers to resolve gene expression patterns with cellular and sub-cellular precision within the complex architecture of developing embryos.
Tissue Preservation Strategies for mRNA Spatial Analysis
Formalin-Fixed Paraffin-Embedding (FFPE) Protocols
Formalin fixation and paraffin embedding represents the gold standard for long-term tissue preservation due to superior morphological preservation and compatibility with clinical biobanking. However, standard formalin fixation introduces mRNA cross-linking and fragmentation, posing significant challenges for spatial transcriptomics. An optimized FFPE protocol for embryonic tissues must balance fixation time to adequately preserve delicate structures while minimizing RNA damage.
Decrosslinking for mRNA Recovery: A critical advancement enabling spatial transcriptomics in FFPE tissues is the implementation of robust decrosslinking procedures. Research indicates that cross-link reversal is optimally performed by heat-induced retrieval at 70°C with Tris-EDTA buffer at pH 8.0 [56]. A pH of 8.0 is crucial as it prevents unwanted side reactions such as pH-dependent RNA hydrolysis. Furthermore, Tris molecules may participate in a quenching mechanism with formaldehyde, enhancing mRNA recovery [56]. Prior to decrosslinking, tissue sections attached to spatially barcoded slides should be deparaffinized by successive immersions in xylene and ethanol, followed by a pre-permeabilization step with collagenase to disrupt extracellular matrix components [56].
Table 1: Optimized Decrosslinking Conditions for FFPE Embryonic Tissues
| Parameter | Recommended Condition | Rationale |
|---|---|---|
| Buffer Composition | Tris-EDTA (TE) | Maintains stable pH environment |
| pH Level | 8.0 | Prevents RNA hydrolysis |
| Temperature | 70°C | Facilitates efficient cross-link reversal |
| Duration | Tissue-dependent optimization | Balances mRNA recovery vs. degradation |
Alternative Preservation Methods
While FFPE dominates clinical pathology, fresh frozen (FF) tissue preservation often yields higher RNA integrity and is preferable when biobanking constraints allow. FF tissues generally maintain higher RNA integrity, but their preparation and storage require stringent conditions [57]. For embryonic research, rapid freezing in optimal cutting temperature (OCT) compound using dry ice or liquid nitrogen-cooled isopentane effectively preserves RNA while maintaining tissue architecture. The choice between FFPE and FF approaches involves weighing priorities: FFPE offers superior morphological detail and stable storage at ambient temperatures, while FF provides higher RNA quality for sensitive transcript detection.
Tissue Sectioning Parameters for Embryonic Tissues
Section Thickness Optimization
Tissue section thickness represents a critical parameter influencing mRNA capture efficiency in spatial transcriptomics. As indicated in recent methodological reviews, "when preparing tissue sections for spatial transcriptomics studies, thickness is critical. If the sections are too thick, they become difficult to penetrate, resulting in greater RNA loss. Conversely, if the sections are too thin, obtaining complete cells becomes challenging" [57]. For most embryonic tissues, a thickness range of 5-10 μm provides an optimal balance, though specific optimization may be required for particularly dense or vacuolated tissues.
Sectioning Techniques to Minimize RNA Degradation
Embryonic tissues often exhibit heterogeneous cellular density and fragile extracellular matrices, requiring modified sectioning approaches. Utilizing high-quality microtome blades, maintaining consistent sectioning speed, and employing adhesive tapes or charged slides can prevent tissue folding and tearing. For cryosectioned samples, maintaining consistent temperature (-18°C to -22°C) throughout sectioning is vital to prevent freeze-thaw artifacts that compromise RNA integrity. Immediate processing or storage at -80°C is mandatory to preserve RNA for subsequent spatial analysis.
Tissue Permeabilization Strategies for mRNA Accessibility
Enzymatic Permeabilization Optimization
Permeabilization enables mRNA release from tissue sections for capture by spatially barcoded oligos. This step must be carefully optimized based on tissue type and fixation method, as "specific features that may affect mRNA accessibility vary across tissues" [56]. For FFPE embryonic tissues, enzymatic permeabilization following decrosslinking is recommended. The optimal permeabilization duration must be determined empirically for each tissue type and embryonic stage through a Tissue Optimization (TO) assay [56]. In mouse brain tissue, for instance, the optimal signal was determined at a permeabilization duration of 30 minutes [56].
Tissue Optimization Assay: A standard TO assay involves applying a fluorescently-labeled oligo(dT) probe to tissue sections after permeabilization, followed by reverse transcription. The resulting fluorescent cDNA signal serves as a proxy for mRNA accessibility. Testing a range of permeabilization times (e.g., 12-30 minutes) helps identify the optimal duration that maximizes signal without causing excessive diffusion of mRNA molecules.
Addressing Permeabilization Challenges in Embryonic Tissues
Embryonic tissues present unique permeabilization challenges due to varying extracellular matrix composition, high cellular density, and stage-specific characteristics. Tissues with abundant mesenchymal components may require combination approaches incorporating both enzymatic and detergent-based permeabilization. For methods utilizing random hexamer primers instead of poly(T) capture, such as Stereo-seq V2, permeabilization must be sufficient to allow access to internal RNA fragments in addition to poly(A) tails [57].
Table 2: Permeabilization Conditions for Different Embryonic Tissue Types
| Tissue Type | Recommended Method | Key Considerations |
|---|---|---|
| Early Embryo (E8-E12) | Mild detergent (0.1%-0.5% Triton X-100) | High nuclear-to-cytoplasmic ratio; fragile epithelia |
| Developing CNS | Combination enzymatic/detergent | Dense neuroepithelium; high lipid content |
| Cartilage Primordia | Collagenase-enhanced enzymatic | Abundant extracellular matrix |
| Developing Heart | Optimized enzymatic permeabilization | High cytoplasmic protein content |
Quality Control Assessment for Spatial Transcriptomics
RNA Quality Metrics and Their Limitations
Traditional RNA quality assessments like RNA Integrity Number (RIN) and DV200 provide preliminary indications of RNA degradation but possess limited predictive power for spatial transcriptomics success. Discrepancies have been observed where "one set of clinical carcinosarcoma samples had relatively low RIN values (observed: 2.3–2.5, preferred: >4.0) but still provided good transcriptome coverage upon sequencing, while another set with similar RIN values rendered sequencing data with high duplicate content" [56]. These metrics, based on ribosomal RNA integrity or total RNA, fail to account for the possible loss of the poly(A) tail of mRNA caused by formaldehyde treatment or spatial variance in mRNA abundance and accessibility [56].
Advanced QC Methods for Spatial Applications
To address limitations of conventional QC methods, a template-switch oligo (TSO)-based quality control assay provides a more reliable assessment of spatial mRNA accessibility [56]. This method uses a fluorescently labeled DNA oligo designed to hybridize with the TSO sequence on surface-bound cDNA, where the fluorescence signal serves as a proxy to detect amplifiable fragments specifically [56]. This approach directly accounts for spatial variance in mRNA abundance, accessibility, and the critical factor of cross-link obstruction that may hinder reverse transcription.
For embryonic tissues specifically, incorporating cytoplasmic stains like RiboSoma in DART-FISH protocols can provide quality control by visualizing cDNA content distribution and confirming retention of nucleic acids through the procedural workflow [58]. This staining "serves as a marker for cDNA content of the tissue and provides a quality control for in situ reactions" [58].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagent Solutions for Embryonic Spatial Transcriptomics
| Reagent/Category | Function | Specific Examples/Notes |
|---|---|---|
| Spatial Barcoded Slides | Capture location-specific mRNA | 10x Genomics Visium slides; Slide-seq chips |
| Decrosslinking Buffers | Reverse formalin-induced crosslinks | Tris-EDTA buffer, pH 8.0; specialized commercial kits |
| Permeabilization Reagents | Enable mRNA release from tissue | Collagenase; Proteinase K; detergents (Triton X-100, SDS) |
| Reverse Transcription Master Mix | Generate cDNA from captured mRNA | Template-switching enzymes; fluorescence-compatible mixes |
| Padlock Probes | Target-specific amplification | Array-synthesized probes for in situ hybridization |
| Polyacrylamide Gel Matrix | Secure cDNA/rolonies in situ | DART-FISH embedding protocol [58] |
| Nuclease-Free Water | Prevent RNA degradation | Essential for all solution preparation |
Visualizing Workflows and Signaling Pathways
Experimental Workflow for FFPE Spatial Transcriptomics
Spatial Transcriptomics FFPE Workflow: This diagram outlines the key stages in processing formalin-fixed paraffin-embedded tissues for spatial transcriptomics analysis, from initial tissue preparation through to sequencing.
mRNA Capture and Detection Pathways
mRNA Capture and Detection Methods: This diagram illustrates the relationship between different probe design strategies, capture technologies, and efficiency enhancement approaches in spatial transcriptomics.
Optimizing tissue preservation, sectioning, and permeabilization for embryonic tissues requires careful consideration of the competing demands of morphological preservation and molecular integrity. The protocols outlined in this technical guide provide a foundation for implementing robust spatial transcriptomics in embryonic research. As spatial technologies continue to evolve, with innovations in nanomaterial-enhanced capture, microfluidic processing, and computational prediction, the potential for resolving gene expression patterns with unprecedented resolution in developing tissues continues to expand. By adhering to these optimized protocols and incorporating appropriate quality control measures, researchers can reliably generate high-quality spatial gene expression data from embryonic tissues, advancing our understanding of developmental processes and their dysregulation in disease states.
Addressing Molecular Diffusion and Enhancing Capture Efficiency
In the field of developmental biology, understanding the spatial expression of mRNA within embryos is crucial for unraveling the complex processes of cell differentiation, tissue patterning, and morphogenesis. A significant technical challenge in this domain involves overcoming molecular diffusion effects and enhancing capture efficiency during spatial transcriptomic analysis. Traditional methods relying on time-intensive imaging specialized equipment have limited the scalability and accessibility of high-resolution spatial mapping in embryonic tissues [8]. This technical guide examines innovative computational and molecular approaches that address these fundamental limitations, enabling researchers to achieve precise spatial gene expression profiling in developing embryonic systems. The ability to accurately capture and localize mRNA transcripts within the intricate architecture of embryos provides unprecedented opportunities to decode the molecular mechanisms driving embryonic development and the etiology of developmental disorders.
Core Challenges in Embryonic mRNA Capture
The accurate detection of mRNA spatial expression in embryos presents several distinct technical hurdles that directly impact data quality and biological interpretation.
Molecular Diffusion Artifacts
In embryonic tissues, mRNA molecules are subject to substantial diffusion effects during sample preparation, particularly during tissue fixation and processing steps. This diffusion can blur the precise subcellular localization of transcripts, leading to inaccurate spatial mapping data. The challenge is especially pronounced in late-stage embryonic tissues where cellular density and extracellular matrix composition create complex microenvironments that influence molecular movement [6]. Without proper containment strategies, diffusion artifacts can obscure critical expression boundaries and gradient patterns that define embryonic organization.
Capture Efficiency Limitations
Conventional spatial transcriptomics methods suffer from limited mRNA capture efficiency, resulting in incomplete transcriptome profiling and reduced sensitivity for detecting low-abundance transcripts. In embryonic research, where tightly regulated genes often exhibit spatially restricted expression patterns, this limitation can lead to false negative results and incomplete molecular characterization of developing structures [59]. The problem is compounded by the small quantities of mRNA present in individual embryonic cells and the technical difficulty of handling miniature tissue structures without damage or loss.
Innovative Approaches and Methodologies
Computational Array Reconstruction
A groundbreaking approach developed by researchers at the Broad Institute eliminates the need for physical imaging altogether, instead using computational methods to reconstruct spatial locations of gene expression. This method builds upon the Slide-seq technique but introduces "transmitter" and "receiver" beads containing DNA barcodes that enable molecular diffusion to become a spatial mapping mechanism rather than a limitation [8].
Mechanism of Action: When exposed to UV light, barcodes cleave from transmitter beads, diffuse through the tissue matrix, and are captured by receiver beads. Critically, receiver beads in closer proximity to transmitters capture more DNA barcodes, creating a diffusion-based distance measurement system. Subsequent sequencing measures the levels of captured barcodes, providing information about both gene expression and spatial bead locations [8].
Implementation Workflow:
- Prepare specialized bead arrays containing transmitter and receiver beads with unique DNA barcodes
- Place embryonic tissue sections on the bead array
- Apply UV light to cleave barcodes from transmitter beads
- Allow controlled diffusion and capture of barcodes by receiver beads
- Sequence captured mRNA and barcode molecules
- Apply the Uniform Manifold Approximation and Projection (UMAP) algorithm to reconstruct original spatial locations based on diffusion patterns
This innovative approach has demonstrated capability to map larger tissue sections (up to 1.2 centimeters wide in mouse embryo tissue) compared to previous methods limited to approximately 3 millimeters, significantly advancing the potential for comprehensive embryonic organ mapping [8].
Direct Planar Capture for Engineered Embryonic Tissues
For embryonic cell cultures and engineered tissues incompatible with standard embedding and sectioning, researchers have developed a direct planar capture method that enhances mRNA retention and localization accuracy. This protocol modifies the Visium HD Spatial Gene Expression workflow to enable compatibility with adherent embryonic cell cultures by growing cells directly on hydrogel-coated microscope slides, eliminating the need for physical sectioning that often exacerbates molecular diffusion [59].
Table 1: Key Reagents for Direct Planar Capture Protocol
| Reagent | Function | Specifications |
|---|---|---|
| Hydrogel-coated microscope slides | Provides substrate for cell growth and mRNA capture | Standard microscope slide format compatible with Visium HD |
| PureCol Type I Bovine Collagen Solution | Surface coating for cell adhesion | Advanced Biomatrix 5005-100ML |
| Paraformaldehyde (16%) | Tissue fixation to minimize diffusion | Electron Microscopy Sciences 15710 |
| Permeabilization reagents | Enable mRNA access for capture | Specific formulation optimized for embryonic cells |
| Visium HD Capture Slide | Barcoded spatial transcriptomics platform | 10x Genomics, with 2μm × 2μm barcoded spots |
Critical Protocol Steps:
- Sterilize microscope slides by submerging in 70% ethanol for 30 minutes
- Coat slides with collagen solution to enhance embryonic cell adhesion
- Culture embryonic cells directly on prepared slides
- Fix cells in place using 4% paraformaldehyde for 24 hours at room temperature
- Permeabilize tissues using optimized conditions for embryonic cells
- Proceed with standard Visium HD spatial transcriptomics workflow
This method preserves the spatial arrangement of embryonic cells without the sample loss or diffusion artifacts associated with physical sectioning, significantly enhancing capture efficiency for delicate embryonic tissues [59].
Advanced Visualization and Single-Molecule Detection
For validation and high-resolution analysis, several advanced visualization techniques provide critical insights into mRNA localization within embryonic structures:
Single-Molecule Fluorescence In Situ Hybridization (smFISH): This method enables detection of individual mRNA molecules within fixed embryonic tissues using multiple fluorescent probes hybridized to single mRNA molecules. Recent advancements allow multiplexed, automated, and high-throughput application to embryonic samples, providing quantitative data on transcript distribution at subcellular resolution [60].
Live-Cell mRNA Imaging Using the MS2 System: For dynamic studies of mRNA localization in living embryonic cells, the MS2 bacteriophage system permits real-time tracking of mRNA movements. This approach involves cloning MS2-binding sites into the mRNA of interest and co-expressing the MS2 coat protein fused to a fluorescent protein. The system has been successfully adapted for embryonic research, revealing mRNA transport mechanisms and localization dynamics during development [60].
Quantitative Performance Data
Table 2: Performance Comparison of Spatial Transcriptomics Methods in Embryonic Research
| Method | Spatial Resolution | Capture Efficiency | Tissue Compatibility | Maximum Tissue Area | Specialized Equipment Needed |
|---|---|---|---|---|---|
| Computational Array Reconstruction [8] | Bead-level (subcellular) | High (utilizes diffusion) | Fresh frozen embryonic tissues | 1.2 cm (demonstrated), 7 cm (in development) | No specialized equipment required |
| Direct Planar Capture [59] | Single-cell (2μm spots) | Standard Visium HD efficiency | Adherent embryonic cell cultures, engineered tissues | Limited by Visium HD capture area | Requires CytAssist instrument |
| Standard Visium HD [6] | 2-10 cells per spot | Standard | FFPE, FxF, FF embryonic tissues | Standard Visium HD slide area | Requires CytAssist instrument |
| smFISH [60] | Single-molecule | Variable (probe-dependent) | Fixed embryonic sections | Unlimited (slide-based) | Standard fluorescence microscopy |
Integration with Embryonic Development Research
The application of these advanced methodologies has yielded significant insights into embryonic development. In late-stage embryonic and postnatal mouse brain research, spatial transcriptomics has enabled the identification of novel molecular markers with precise spatial localization, including Car12 and Folr1 in the choroid plexus, Rprml in the piriform cortex, and Hsd11b2 in the thalamus [6]. These discoveries were facilitated by computational integration of spatial transcriptome data with single-cell RNA-seq datasets, allowing resolution of embryonic brain microstructure despite the challenges of molecular diffusion.
The spatiotemporal analysis of embryonic brain development revealed both conserved and stage-specific gene expression patterns, with distinct transcriptional signatures characterizing progenitor zones in embryonic stages that disappear in adult structures [6]. Such findings highlight the critical importance of precise spatial mapping during embryonic development and the value of technologies that minimize diffusion artifacts while maximizing capture efficiency.
Experimental Workflow Visualization
Essential Research Reagent Solutions
Table 3: Critical Reagents for Advanced Embryonic Spatial Transcriptomics
| Reagent Category | Specific Products | Application in Embryonic Research |
|---|---|---|
| Bead-Based Capture Systems | Slide-seq beads, Visium HD capture slides | Spatial barcoding of embryonic mRNA transcripts |
| Tissue Preservation Reagents | Paraformaldehyde (16%, EM grade), Methanol | Embryonic tissue fixation to minimize diffusion |
| Permeabilization Solutions | SSC Buffer, PureCol Collagen | Enable mRNA accessibility in dense embryonic tissues |
| Library Preparation Kits | Visium HD Spatial Gene Expression kit | Convert captured mRNA to sequencing libraries |
| Computational Tools | Space Ranger, Seurat, UMAP algorithms | Spatial data analysis and visualization for embryonic datasets |
| Quality Control Reagents | KAPA SYBR FAST qPCR Master Mix | Assess library quality and capture efficiency |
The evolving methodologies for addressing molecular diffusion and enhancing capture efficiency in embryonic spatial transcriptomics represent a paradigm shift in developmental biology research. The integration of computational reconstruction approaches with advanced molecular capture techniques has successfully transformed diffusion from a technical obstacle into a measurable spatial parameter. As these technologies continue to mature, researchers are poised to unravel the complex spatial gene expression patterns that orchestrate embryonic development with unprecedented resolution and accuracy. The ongoing development of even larger-scale mapping capabilities—potentially enabling comprehensive analysis of entire embryonic organs—will further accelerate discoveries in fundamental developmental processes and developmental disorders.
The pursuit of understanding mRNA spatial expression in embryonic development presents a unique challenge at the intersection of biological inquiry and fiscal practicality. This technical guide examines the critical balance between sequencing depth and experimental cost, providing researchers with strategic frameworks for designing spatially-resolved transcriptomics studies. By synthesizing current methodologies from bulk RNA-seq to advanced spatial transcriptomics platforms, we delineate how informed decisions on library preparation, sequencing platforms, and multiplexing strategies can optimize both data quality and resource allocation. Within the context of embryonic research, where spatial context is paramount for unraveling developmental mechanisms, these cost-benefit analyses become particularly vital for constructing robust, reproducible experimental pipelines that advance our understanding of patterning and cell fate decisions.
The emergence of sophisticated spatial transcriptomics (sST) technologies has revolutionized mRNA expression analysis in embryonic research by preserving the anatomical context essential for understanding developmental processes. Traditional single-cell RNA sequencing (scRNA-seq), while powerful for dissecting cellular heterogeneity, fundamentally destroys spatial information by requiring tissue dissociation and cell isolation [61]. This limitation is particularly problematic in embryology, where the positional context of gene expression drives pattern formation and cell fate specification. However, this preserved spatial resolution comes with substantial financial considerations, creating a cost-resolution paradox that researchers must navigate.
Sequencing-based spatial transcriptomics (sST) has catalyzed major advancements by facilitating transcriptome-scale spatial gene expression measurement, yet the field lacks comprehensive benchmarking, and variability across platforms presents significant challenges in standardized evaluation and cost estimation [62]. The financial investment required for spatial transcriptomics is substantially higher than conventional bulk RNA-seq, with costs influenced by multiple factors including spatial resolution, capture efficiency, sequencing depth, and data analysis requirements. For researchers investigating mRNA spatial expression in embryos, where sample numbers can be large due to developmental time series or multiple genetic conditions, effective experimental design becomes paramount for balancing budgetary constraints with scientific rigor.
Quantitative Landscape of RNA-Sequencing Costs
Understanding the precise cost breakdown of RNA sequencing is fundamental to effective experimental design. While spatial transcriptomics platforms have their own pricing structures, their underlying chemistry often builds upon conventional RNA-seq workflows, making standard RNA-seq cost drivers highly relevant for budget planning.
Primary Cost Components in RNA-Seq Workflows
A comprehensive mRNA-seq experiment includes multiple cost components that collectively determine the total investment required. Recent analyses indicate that the cost for RNA-sequencing ranges from approximately $36.9 to $173 per sample for a single sample in an mRNA-seq experiment, with variability depending primarily on library preparation methods and sequencing depth [63]. The table below provides a detailed breakdown of these costs across different experimental stages.
Table 1: Cost Breakdown for mRNA-seq Experiments (per sample)
| Cost Category | Specific Method | Cost Range (USD) | Key Parameters |
|---|---|---|---|
| RNA Extraction | TRIzol method | $2.2 | Solvent-based approach [63] |
| QIAgen RNeasy Kit | $7.1 | Silica-based column [63] | |
| Quality Control | Bioanalyzer RNA-6000-Nano | $4.1 | RNA quality assessment [63] |
| Library Preparation | Illumina TruSeq mRNA stranded | $64.4 | Traditional high-quality standard [63] |
| NEBnext Ultra II RNA | $37 | Cost-effective alternative [63] | |
| Lexogen QuantSeq-Pool | $39.5 | 3' mRNA-seq barcoding [63] | |
| Alithea MERCURIUS BRB-seq | $19.7 | Ultra-cost-effective via early pooling [63] | |
| Library QC | Bioanalyzer DNA-1000 | $4.3 | Library quality assessment [63] |
| Sequencing | NovaSeq S4 flow cell (TruSeq) | $36.9 | ≥25M reads/sample at full capacity [63] |
| NovaSeq S4 flow cell (BRB-seq) | $4.6 | 5M reads/sample at full capacity [63] | |
| Data Analysis | Cloud-based pipelines | ~$2 | Plus ongoing storage costs [63] |
Notably, library preparation has emerged as the most expensive step in many RNA-seq experiments, particularly when using traditional kits such as Illumina's TruSeq mRNA stranded library prep at $64.4 per sample [63]. However, technologies employing early barcoding and pooling strategies, such as Bulk RNA Barcoding and sequencing (BRB-seq), dramatically reduce these costs to approximately $19.7 per sample by processing multiple samples in a single library preparation reaction [63].
Sequencing Depth and Platform Considerations
Sequencing costs vary significantly based on the platform and depth required. For the Illumina NovaSeq 6000 system, using the high-capacity S4 300 cycle flow cell represents the most cost-effective option for high-throughput studies, with costs reaching approximately $36.9 per sample when multiplexing around 400 samples for ≥25 million reads per sample [63]. The required sequencing depth depends largely on the library preparation method and experimental goals. While traditional mRNA-seq libraries such as TruSeq typically require 25 million reads or more per sample for robust detection of expressed transcripts, 3' mRNA-seq methods such as BRB-seq can achieve comparable results with only 5 million reads per sample, substantially reducing sequencing costs [63].
Table 2: Sequencing Cost Comparison on NovaSeq Platforms
| Library Method | Reads/Sample | Samples/Flow Cell | Cost/Sample | Best Use Cases |
|---|---|---|---|---|
| Illumina TruSeq | ≥25M | ~400 | $36.9 | Traditional standard, full transcript coverage [63] |
| NEBnext Ultra II | 20M | ~400 | $25.9 | Balanced cost and coverage [63] |
| Lexogen QuantSeq-Pool | 5M | ~3,200 | $4.6 | Cost-sensitive large studies, 3' digital gene expression [63] |
| Alithea BRB-seq | 5M | ~3,200 | $4.6 | Ultra-high throughput, counting-based applications [63] |
Sequencing Depth Requirements for Spatial Expression Analysis
Determining appropriate sequencing depth is particularly crucial for spatial transcriptomics studies in embryonic systems, where capturing both abundant and rare transcripts is essential for comprehending developmental patterning.
Methodological Considerations for Spatial Resolution
Sequencing-based spatial transcriptomics (sST) methods have diversified considerably, with systematic comparisons revealing substantial variability in performance characteristics. A comprehensive benchmarking study evaluating 11 sST methods across reference tissues including mouse embryonic eyes and brain regions revealed that methods differ significantly in spatial resolution, capture efficiency, and molecular diffusion [62]. These technical parameters directly influence the sequencing depth required to achieve satisfactory results.
The resolution of sST methods is typically defined by the distance between spot centers, which ranges from less than 10μm for high-resolution platforms such as Stereo-seq, BMKMANU S1000, and Salus, to 100μm for methods such as DBiT-seq [62]. This spatial resolution directly impacts the number of transcriptome profiles needed to cover an embryo section and consequently influences sequencing requirements. Higher resolution methods generate substantially more spatial data points, potentially requiring greater overall sequencing depth to maintain sufficient coverage per location.
Capture Efficiency and Sensitivity Across Platforms
Sensitivity varies considerably across sST platforms, influencing depth requirements. Benchmarking studies demonstrate that when sequencing depth is controlled through downsampling, Slide-seq V2 shows higher sensitivity in certain tissues such as mouse eyes, while probe-based Visium and DynaSpatial perform better in other contexts such as hippocampus [62]. This variability suggests that optimal platform selection depends on both the tissue type and the specific genes of interest.
Notably, sequencing saturation analyses reveal that typical sST sequencing runs—ranging from 300 million reads for Visium to 4 billion reads for Stereo-seq—often do not reach saturation, indicating that additional sequencing depth could continue to improve sensitivity [62]. This finding highlights the trade-off between data quality and cost, particularly for methods with higher spot densities.
Strategic Experimental Design for Embryonic mRNA Localization
Embryonic systems present unique challenges for spatial transcriptomics, including low mRNA abundance in early development and dynamic expression patterns that require careful temporal sampling. Strategic experimental design must address these challenges while maintaining fiscal responsibility.
Integrating Spatial and Temporal Dimensions
The study of embryonic development necessitates both spatial and temporal resolution. Traditional approaches have faced a fundamental trade-off between anatomical context and quantitation, typically requiring destruction of sample morphology through microdissection, cell dissociation, homogenization, or sorting [64]. Modern sST methods overcome this limitation but introduce cost considerations.
For studies focusing on maternal mRNA localization and inheritance, such as investigations into early zebrafish embryogenesis, enhanced tomo-seq methods provide high spatial resolution along defined axes [32]. In such applications, embedding and orienting individual embryos at specific developmental stages along visible axes (e.g., animal-vegetal) followed by serial sectioning enables transcriptome-wide localization analysis with spatial precision [32]. The financial considerations for these experiments include not only sequencing costs but also sample preparation time and specialized expertise.
Quantitative Imaging-Based Approaches
For laboratories with imaging expertise and access to advanced microscopy, quantitative in situ methods offer an alternative pathway for spatial expression analysis. Techniques such as quantitative Hybridization Chain Reaction (qHCR) enable accurate and precise relative quantitation of mRNA expression with subcellular resolution within whole-mount vertebrate embryos, preserving anatomical context while providing quantitative data [64]. This approach is particularly valuable for validating findings from sequencing-based spatial transcriptomics or for focused studies of smaller gene panels.
The cost structure for imaging-based approaches differs significantly from sequencing-based methods, with higher initial investment in instrumentation but potentially lower per-sample costs for studies with limited gene targets. These methods also enable bi-directional queries: "read-out" from anatomical space to expression space reveals co-expression relationships in selected regions, while "read-in" from expression space to anatomical space reveals locations where specific gene co-expression relationships occur [64].
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Research Reagent Solutions for Spatial Transcriptomics in Embryonic Research
| Category | Specific Product/Platform | Key Function | Application Notes |
|---|---|---|---|
| Library Prep Kits | Illumina TruSeq mRNA stranded | Full-length transcript library prep | High quality but premium cost [63] |
| NEBnext Ultra II RNA | Cost-effective library preparation | Balanced performance for bulk RNA-seq [63] | |
| Alithea MERCURIUS BRB-seq | Ultra-affordable library prep | Early barcoding and pooling for cost reduction [63] | |
| Spatial Platforms | 10X Genomics Visium | Array-based spatial transcriptomics | Probe and polyA-based versions available [62] |
| Stereo-seq | High-resolution spatial transcriptomics | Small distance between spot centers (<10μm) [62] | |
| Slide-seq V2 | Bead-based spatial transcriptomics | High sensitivity in certain tissues [62] | |
| DBiT-seq | Microfluidics-based spatial mapping | 100μm resolution, compatible with small embryos [62] | |
| In Situ Technologies | qHCR (Quantitative HCR) | Quantitative in situ hybridization | Subcellular resolution in whole-mount embryos [64] |
| smFISH | Single-molecule mRNA counting | Absolute transcript counts with spatial context [65] | |
| Specialized Methods | Tomo-seq | Spatial transcriptomics of sections | Ideal for axial patterning studies [32] |
| scSLAM-seq | Single-cell metabolic labeling | Distinguishes maternal and zygotic transcripts [32] |
Decision Framework and Workflow Integration
Navigating the complex landscape of methodological options requires a systematic approach to experimental design. The following workflow provides a logical decision framework for researchers planning studies of mRNA spatial expression in embryos.
Diagram 1: Experimental Design Workflow for Spatial Embryonic Studies
The investigation of mRNA spatial expression in embryos demands careful consideration of the interplay between methodological resolution, biological questions, and fiscal constraints. No single approach optimally addresses all experimental scenarios—rather, the selection of platforms and parameters must align with specific research goals, tissue characteristics, and analytical requirements. As spatial technologies continue to evolve, with improvements in resolution, sensitivity, and multiplexing capabilities, the cost-benefit calculus will undoubtedly shift. Nevertheless, the fundamental principles outlined in this guide—strategic technology selection, appropriate depth determination, and systematic workflow optimization—provide a enduring framework for designing spatially-resolved transcriptomics studies that maximize scientific insight while maintaining fiscal responsibility. Through deliberate experimental planning that balances these competing priorities, researchers can effectively unravel the complex spatial dynamics of gene expression that underlie embryonic development.
Computational Solutions for Improving Data Decoding and Resolution
The precise detection of mRNA spatial expression in embryos is fundamental to understanding the complex processes of developmental biology, including lineage specification and cellular differentiation [66]. Spatially resolved transcriptomic technologies have emerged as pivotal tools for this purpose, enabling the profiling of gene expression throughout tissues while retaining crucial spatial information about cellular original locations [67]. However, these technologies present a significant trade-off: imaging-based methods offer high spatial resolution but limited transcriptome coverage, while next-generation sequencing (NGS)-based approaches provide extensive transcriptome profiling but often at reduced spatial precision where capture spots may contain mixed transcripts from multiple cells [67] [68].
This resolution limitation is particularly pronounced in embryonic research, where the dynamic and regulative nature of early embryogenesis introduces intrinsic variation into each dataset [66]. The intricate spatial architecture of embryonic tissues, combined with the technical challenges of working with limited cellular material, necessitates advanced computational approaches to deconvolve mixed cellular signals and reconstruct high-resolution spatial expression maps [67] [66]. This whitepaper examines current computational methodologies designed to overcome these limitations, with specific application to embryonic development research.
Core Computational Methodologies
Algorithmic Approaches for Spatial Deconvolution
Multiple computational strategies have been developed to address the challenge of cell-type deconvolution in spatial transcriptomics data. These approaches can be broadly classified into several categories based on their underlying mathematical principles and processing paradigms [67].
Table 1: Categories of Computational Deconvolution Approaches
| Approach Category | Core Principle | Key Strengths | Example Algorithms |
|---|---|---|---|
| Probabilistic Models | Uses statistical distributions (Negative Binomial, Poisson) to model gene expression and infer cell-type proportions [67]. | Effectively handles technical noise and count-based nature of transcriptomic data. | Cell2location [67], DestVI [67], RCTD [67], Stereoscope [67] |
| Non-negative Matrix Factorization (NMF) | Factorizes gene expression matrix into non-negative components representing cell-type signatures and proportions [67]. | Uncover latent patterns without requiring reference data. | SPOTlight [67], SpatialDWLS [67], NMFreg [67] |
| Deep Learning Frameworks | Employs neural networks to integrate datasets and learn complex patterns for cell type classification [66]. | Capable of modeling non-linear relationships in large, complex datasets. | scVI [66], scANVI [66], scGen [66] |
| Graph-Based Methods | Utilizes graph theory to model spatial relationships between spots or cells for informed deconvolution [67]. | Incorporates spatial context directly into the modeling process. | GraphST [68], DSTG [67] |
| Optimal Transport Theory | Applies mathematical frameworks for transferring mass between distributions to map cells to spatial locations. | Effective for aligning single-cell and spatial datasets. | CytoSPACE [68] |
Integrated Mapping Approaches
Beyond deconvolution, newer methodologies focus on precisely mapping single cells to their spatial contexts. The Cellular Mapping of Attributes with Position (CMAP) algorithm exemplifies this approach through a sophisticated three-level process [68]:
- CMAP-DomainDivision: Partitions cells into spatial domains using hidden Markov random field (HMRF) and spatially specific genes, then trains a classification model (e.g., Support Vector Machine) to assign spatial domain labels to individual cells [68].
- CMAP-OptimalSpot: Identifies spatially variable genes within each domain, generates a random alignment matrix between cells and spots, then employs deep learning-based optimization with a Structural Similarity Index (SSIM) metric to arrive at an optimal mapping matrix [68].
- CMAP-PreciseLocation: Builds a nearest neighbor graph representing spot relationships, then uses a Spring Steady-State Model learned from physical field to assign exact (x, y) coordinates to each cell [68].
This integrated approach has demonstrated superior performance in benchmarking studies, achieving a 73% weighted accuracy in mapping cells to correct spots while maintaining a 99% cell usage ratio in simulated mouse olfactory bulb data [68].
Experimental Protocols and Methodologies
Reference Model Construction for Embryonic Development
For embryonic research applications, constructing a high-quality reference model requires careful data collection and processing:
Data Collection Criteria:
- Select only peer-reviewed published datasets with complete cell metadata for collection time and cell type annotations [66].
- Filter to include only wild-type embryos, excluding experimental genotypes or in vitro stem cells [66].
- Implement automated preprocessing pipelines (e.g., nf-core) to ensure consistency in alignment, quantification, and utilization of updated genome assemblies [66].
Species-Specific Processing:
- For mouse embryos: Normalize SMART-seq1/2 datasets by gene length, discard ribosomal mitochondrial genes, and retain cells with minimum of 20,000 transcripts per cell [66].
- For human embryos: Address ambiguous cell labels by setting them to 'Unknown', collapse early stages (before 8-cell) to 'prelineage' categories to bolster cell numbers, and transform read counts by gene length for UMI-based technology compatibility [66].
Integration and Validation:
- Employ deep learning integration techniques (scVI, scANVI) with fine-tuned parameters using autotune features in scvi-tools [66].
- Validate models through downstream analyses including nearest-neighbor graph computation, force-directed graph layout, UMAP dimension reduction, and unsupervised Leiden clustering [66].
- Infer differentiation trajectories using partition-based graph abstraction (PAGA) to verify biological relevance [66].
Workflow Visualization
Workflow for Spatial mRNA Analysis in Embryos
CMAP Three-Level Mapping Process
Performance Benchmarking and Validation
Quantitative Assessment of Algorithm Performance
Rigorous benchmarking is essential for evaluating computational deconvolution and mapping methods. Performance assessment should include multiple metrics to capture different aspects of accuracy and utility.
Table 2: Performance Comparison of Spatial Mapping Algorithms
| Algorithm | Mapping Accuracy | Cell Usage Ratio | Computational Efficiency | Reference Requirement |
|---|---|---|---|---|
| CMAP | 73% (weighted accuracy) [68] | 99% (2215/2242 cells) [68] | Moderate (divide-and-conquer strategy) [68] | Optional (works with or without reference) [68] |
| CellTrek | Lower than CMAP [68] | 45% cell loss ratio (1243/2242 unmapped) [68] | Moderate | Yes [68] |
| CytoSPACE | Lower than CMAP [68] | 48% cell loss ratio (1078/2242 unmapped) [68] | Moderate | Yes (for cell number estimation) [68] |
| Cell2location | High-resolution mapping capability [67] | Estimates absolute abundances [67] | High (integrated multi-dataset analysis) [67] | Yes [67] |
| DestVI | Multi-resolution deconvolution [67] | Joint modeling of single-cell and spatial data [67] | High (automated downstream pipeline) [67] | Yes [67] |
Method Selection Guidelines
Choosing the appropriate computational method depends on specific experimental conditions and research objectives:
- When comprehensive reference data is available: Probabilistic models like Cell2location and DestVI provide robust deconvolution with explicit uncertainty quantification [67].
- When reference data is limited or unavailable: Reference-free approaches like STdeconvolve (LDA-based) and CARD (reference-free capability) enable cell type discovery directly from spatial data [67].
- For maximum spatial precision: Integrated mapping tools like CMAP offer superior coordinate-level assignment, particularly valuable for studying fine-scale cellular interactions in embryonic development [68].
- For large-scale dataset integration: Deep learning frameworks like scVI and scANVI effectively model non-linear relationships and batch effects across multiple embryonic datasets [66].
Research Reagent Solutions
Table 3: Essential Research Reagents and Platforms for Spatial Embryonic Analysis
| Reagent/Platform | Function | Application in Embryonic Research |
|---|---|---|
| 10x Genomics Visium | NGS-based spatial transcriptomics with spot resolution [67] | Mapping gene expression in embryonic sections with transcriptome-wide coverage [67] |
| Slide-seq | High-density spatial transcriptomics using barcoded microbeads [67] | Near-single-cell resolution mapping of embryonic tissues [67] |
| Seq-Scope | Illumina flow cell-based patterned oligonucleotide arrays [67] | Submicron-scale mRNA measurement in embryonic structures [67] |
| Stereo-Seq | DNA nanoball (DNB) arrays for nanometer precision [67] | High-resolution assessment across extensive embryonic areas [67] |
| scVI-tools | Probabilistic modeling package for single-cell omics data [66] | Integration and classification of multiple embryonic datasets [66] |
| CARD Framework | Spatially aware deconvolution with reference-free capability [67] | Reconstruction of high-resolution expression in embryonic tissues [67] |
Computational solutions for improving data decoding and resolution have become indispensable tools in the study of mRNA spatial expression in embryos. These methodologies directly address the fundamental trade-offs in spatial transcriptomics technologies, enabling researchers to extract maximum biological insight from precious embryonic materials. The continuing development of probabilistic models, deep learning frameworks, and integrated mapping approaches like CMAP promises to further enhance our ability to resolve the complex spatial architecture of developing embryos at single-cell resolution. As these computational methods evolve, they will increasingly enable the precise delineation of lineage specification events, cellular differentiation pathways, and spatial organization principles that underlie embryonic development.
Benchmarking Performance and Ensuring Biological Relevance
The precise spatial organization of gene expression is a fundamental regulator of embryonic development, directing processes like cell fate determination, tissue patterning, and organogenesis. Spatial transcriptomics (ST) has emerged as a pivotal technology that bridges the critical gap between single-cell RNA sequencing (scRNA-seq) and tissue morphology by mapping gene expression data directly onto its original histological context [69] [61]. For embryonic research, where cellular location is intrinsically linked to function and developmental trajectory, this spatial context is indispensable. The technology can be broadly categorized into two groups: sequencing-based spatial transcriptomics (sST), which uses spatially barcoded arrays and next-generation sequencing, and imaging-based spatial transcriptomics (iST), which relies on multiplexed fluorescence in situ hybridization to localize transcripts [3] [70]. As the commercial ST landscape rapidly evolves, offering platforms with subcellular resolution and expanded gene panels, systematic benchmarking of their sensitivity, resolution, and control over molecular diffusion is essential for researchers to select the optimal tool for investigating mRNA spatial expression in embryos [69] [62].
Platform Categorization and Core Technologies
Imaging-Based Spatial Transcriptomics (iST)
iST platforms utilize iterative hybridization, imaging, and dye-removal cycles to decode the spatial locations of RNA transcripts. They offer single-molecule resolution but are typically limited to targeted gene panels [3].
- Xenium (10x Genomics): This platform employs a padlock probe method. Probes hybridize to target RNA and are circularized and amplified via rolling circle amplification (RCA) to enhance signal. Fluorescently labeled probes then bind to these rolling circle products, and their cyclic imaging generates a unique optical signature for each gene [3] [70].
- MERFISH (Vizgen): MERFISH uses a binary barcoding strategy. Each gene is assigned a unique barcode of '0's and '1's. Through multiple rounds of hybridization and imaging, the presence ('1') or absence ('0') of fluorescence is recorded to build the barcode for each RNA molecule, enabling error-resistant identification [3].
- CosMx (NanoString): CosMx combines elements of both hybridization and optical signature methods. It uses probes with a readout domain containing multiple sub-domains. The binding of fluorescent secondary probes to these sub-domains over multiple cycles generates a unique combination of color and position for each target gene, allowing for a high degree of multiplexing [3].
Sequencing-Based Spatial Transcriptomics (sST)
sST platforms capture mRNA onto surfaces coated with spatially barcoded oligonucleotides. The captured transcripts are then sequenced, and their locations are inferred from the spatial barcodes [3].
- Visium HD (10x Genomics): Building on the original Visium technology, Visium HD uses a pair of probes that hybridize to the target mRNA and are then ligated. This ligated product is captured by poly(dT) on the array. The key advancement is a significantly reduced spot size of 2 μm, enabling subcellular resolution [69] [3].
- Stereo-seq (BGI): This platform uses DNA Nanoball (DNB) technology. Oligo probes containing a poly(dT) sequence and spatial barcodes are circularized and amplified into DNBs, which are then patterned onto a chip at very high density. With a center-to-center distance of just 0.5 μm, Stereo-seq offers one of the highest spatial resolutions among commercial sST platforms [69] [3].
Table 1: Core Technological Characteristics of Major Spatial Transcriptomics Platforms.
| Platform | Technology Type | Key Technology | Resolution (Spot/Center Distance) | Transcriptome Coverage |
|---|---|---|---|---|
| Xenium | Imaging-based (iST) | Padlock Probes & RCA | Subcellular | Targeted (~5000 genes) |
| MERFISH | Imaging-based (iST) | Binary Barcoding | Single-molecule | Targeted (500-1000 genes) |
| CosMx | Imaging-based (iST) | Hybridization & Cyclic Imaging | Single-cell to subcellular | Targeted (1000-6000 genes) |
| Visium HD | Sequencing-based (sST) | Spatial Barcoding on Array | 2 μm spot size | Whole Transcriptome (18,000 genes) |
| Stereo-seq | Sequencing-based (sST) | DNA Nanoball (DNB) Array | 0.5 μm center-to-center | Whole Transcriptome |
Figure 1: Spatial transcriptomics platforms are broadly categorized into imaging-based (iST) and sequencing-based (sST) technologies, each with distinct chemistries that collectively determine key performance metrics like sensitivity, resolution, and diffusion control.
Systematic Benchmarking of Performance Metrics
Sensitivity and Molecular Capture Efficiency
Sensitivity, or the ability to detect low-abundance transcripts, is critical in embryonic studies where key morphogens may be expressed at low levels. Systematic evaluations using matched tissue sections reveal significant differences between platforms.
A comprehensive benchmark of high-throughput platforms using colon adenocarcinoma, hepatocellular carcinoma, and ovarian cancer samples found that Xenium 5K consistently demonstrated superior sensitivity for multiple marker genes, even when analysis was restricted to shared tissue regions to reduce bias [69]. In the same study, when assessing the entire gene panel, Stereo-seq, Visium HD, and Xenium 5K showed high gene-wise correlation with matched scRNA-seq data, indicating their robust capture efficiency. In contrast, CosMx 6K, while sometimes detecting a higher total number of transcripts, showed substantial deviation from scRNA-seq reference data, a discrepancy not fully resolved by stricter quality control thresholds [69].
Another systematic comparison of sST platforms using mouse embryonic eyes highlighted the impact of methodology and sequencing depth. Probe-based Visium and DynaSpatial showed high sensitivity for marker genes in the lens and neuronal retina. Notably, Slide-seq V2 also demonstrated high sensitivity when sequencing depth was controlled for, whereas polyA-based Visium did not generate as many counts for expected marker genes [62]. This underscores that capture chemistry, not just raw sequencing depth, is a major determinant of sensitivity.
Spatial Resolution and Molecular Diffusion
The "effective resolution" of a platform is a function of its nominal spot size or probe size and its control over molecular diffusion during tissue processing. Diffusion can cause mRNA molecules to migrate from their original location, blurring the spatial data. This is particularly problematic in embryonic tissues, which can be rich in RNases and require specific handling.
sST platforms exhibit a wide range of nominal resolutions. Stereo-seq offers the highest density with a 0.5 μm center-to-center distance between its DNA nanoballs, while Visium HD uses a 2 μm spot size [69] [3]. However, a benchmark of 11 sST methods identified molecular diffusion as a critical and variable parameter that significantly affects this nominal resolution, with diffusion rates varying across different methods and tissue types [62]. This means the effective resolution can be lower than the physical spot size.
iST platforms, by contrast, generally offer superior diffusion control and single-molecule resolution. Xenium, in particular, has been noted for its enhanced diffusion control and minimized spatial artifacts, contributing to high spatial precision [69]. This makes iST platforms particularly suited for mapping fine-grained spatial patterns, such as those in developing embryonic structures.
Table 2: Benchmarking Performance Metrics Across Spatial Transcriptomics Platforms.
| Platform | Sensitivity (Marker Genes) | Correlation with scRNA-seq | Effective Resolution | Key Finding from Benchmarking |
|---|---|---|---|---|
| Xenium | High [69] | High [69] [70] | Single-cell / Subcellular [69] | Superior sensitivity & diffusion control; precise cell typing [69] [71] |
| CosMx | Variable [69] | Lower in some studies [69] | Single-cell / Subcellular [72] | High total transcripts but may deviate from scRNA-seq [69] |
| MERFISH | Information Missing | Information Missing | Single-molecule [3] | Performance can be tissue-quality dependent [72] |
| Visium HD | High [69] | High [69] | ~8 μm (after binning) [69] | Robust, transcriptome-wide maps; good concordance with CODEX [69] |
| Stereo-seq | High [69] [62] | High [69] | Subcellular [69] | Highest capture capability & sequencing reads requirement [69] [62] |
Figure 2: The workflow from tissue preparation to data output shows that mRNA diffusion during permeabilization is a major factor determining a platform's effective resolution. iST platforms typically exert better diffusion control, leading to higher effective resolution.
Application to Embryonic Development Research
Experimental Design and Protocol for Embryonic Studies
To ensure reliable results in embryonic research, a rigorous experimental design is paramount. The following protocol, synthesized from recent benchmarking studies, outlines key steps:
- Sample Preparation and Matching: For a fair platform comparison or integrative analysis, use serial tissue sections from the same embryo block. This controls for biological variability. For FFPE-embedded samples, ensure uniform baking times after slicing to standardize tissue adherence and RNA retention across all sections destined for different platforms [70].
- Ground Truth Establishment: Generate orthogonal datasets from the same sample to validate ST results.
- Perform single-cell RNA sequencing (scRNA-seq) on a dissociated portion of the matched sample to provide a reference transcriptome for cell type annotation and correlation analysis [69] [70].
- Use multiplex immunofluorescence (e.g., CODEX) or H&E staining on adjacent serial sections to obtain high-resolution protein and morphological data for spatial validation [69] [72].
- Data Processing and Normalization: To compare sensitivity across platforms, normalize for varying sequencing depths. This can be done by downsampling all datasets to an equivalent number of reads before comparing transcript counts within predefined anatomical regions, such as the lens or neuronal retina of an embryonic mouse eye [62].
- Region of Interest (ROI) Analysis: Manually delineate regions based on clear histological architecture (e.g., embryonic brain layers, organ primordia). Analyzing performance within these shared ROIs minimizes variability introduced by analyzing different tissue areas [62].
Insights from Embryonic Tissue Benchmarks
Benchmarking studies using embryonic mouse tissues have yielded critical insights. The mouse embryonic eye, with its well-defined structure of a lens surrounded by neuronal retina cells, serves as an ideal model for evaluating platform performance [62]. Studies using this model have shown that certain platforms, like probe-based Visium and Slide-seq V2, can effectively capture known marker genes in these structures, while others may miss key transcripts, highlighting the need for careful platform selection based on the specific genes of interest [62].
Furthermore, the expansive gene panels of newer platforms (e.g., CosMx 6K, Xenium 5K) are particularly advantageous for embryonic research. The substantially expanded gene panels enable enhanced resolution of cellular states and broader coverage of signaling pathway activities, which is crucial for understanding complex developmental processes [69].
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Key Research Reagent Solutions for Spatial Transcriptomics.
| Item / Reagent | Function in Workflow | Application Note |
|---|---|---|
| Formalin-Fixed Paraffin-Embedded (FFPE) Tissue | Standard for sample preservation; maintains morphology and allows use of archival biobanks. | Essential for clinical & translational research; requires protocols optimized for degraded RNA [72] [70]. |
| Optimal Cutting Temperature (OCT) Compound | Medium for embedding fresh-frozen tissues. | Preserves RNA integrity better than FFPE for some applications; standard for some sST platforms [69]. |
| CytAssist Instrument (10x Genomics) | Facilitates probe transfer from standard slides to Visium slide for FFPE samples. | Simplifies workflow for Visium HD FFPE assays [3]. |
| CODEX Multiplex Immunofluorescence | Provides high-plex protein expression data from adjacent serial sections. | Used for orthogonal validation of transcriptional data with protein-level spatial information [69]. |
| Negative Control & Blank Probes | Assess background noise and signal specificity in iST data. | Critical for quality control; numbers and types vary by platform (e.g., CosMx has 10 negative controls) [72]. |
| DAPI Stain | Fluorescent stain for cell nuclei. | Used for automated and manual cell segmentation across all major platforms [69]. |
The choice of an optimal spatial transcriptomics platform for embryonic research involves balancing multiple factors, including sensitivity, resolution, transcriptome coverage, and compatibility with sample type.
For studies requiring the highest possible sensitivity for a targeted gene panel and superior single-cell resolution, imaging-based platforms like Xenium are a strong choice, as they consistently demonstrate high sensitivity and excellent diffusion control [69] [70]. When the research question demands unbiased, whole-transcriptome discovery without a pre-defined gene panel, sequencing-based platforms like Visium HD and Stereo-seq are more appropriate. Visium HD provides robust, high-resolution maps with good concordance to orthogonal data, while Stereo-seq offers the highest nominal resolution for sST [69] [62].
Ultimately, the decision should be guided by the specific biological question, the required resolution to discern embryonic structures of interest, the importance of detecting low-abundance transcripts, and the availability of high-quality, often precious, embryonic tissue samples. As these technologies continue to advance, the research community will benefit from ongoing, systematic benchmarks that further refine our understanding of their capabilities and optimal applications in developmental biology.
Integration with scRNA-seq Data for Cell Type Deconvolution and Validation
The integration of single-cell RNA sequencing (scRNA-seq) data with spatial transcriptomics (ST) technologies has emerged as a powerful computational approach for elucidating cellular heterogeneity within a spatial context. This technical guide provides a comprehensive overview of methodologies, tools, and experimental protocols for deconvoluting cell types and validating spatial architecture, with particular emphasis on applications in embryonic development research. By synthesizing transcriptomic signatures from dissociated single cells with spatial positioning data, researchers can reconstruct tissue environments at single-cell resolution, enabling unprecedented investigation of spatial gene expression patterns, cell fate determination, and morphogenetic processes in developing embryos.
Cell type deconvolution represents a class of computational methods that infer cellular composition from complex tissue samples by integrating scRNA-seq reference data with spatial transcriptomic profiles. In embryonic research, where spatial organization drives developmental processes, these techniques are particularly valuable for mapping the precise locations of cell types and states without requiring specialized spatial imaging equipment [8]. The fundamental challenge addressed by deconvolution methods is the limitation of most spatial transcriptomics technologies—either high spatial resolution with limited transcriptome coverage (image-based approaches) or comprehensive transcriptome coverage with low spatial resolution (sequencing-based approaches) [73].
Spatial transcriptomics technologies have revolutionized our ability to investigate gene expression patterns while preserving spatial context, but they face inherent limitations. Sequencing-based approaches like 10x Visium, Slide-seq, and Stereo-seq can profile transcriptome-wide gene expression within spatial spots but often contain multiple cells per spot, limiting cellular resolution [73]. Image-based approaches such as MERFISH and seqFISH provide single-cell resolution but typically measure only hundreds to thousands of pre-selected genes, limiting incidental discoveries [73]. Deconvolution methods bridge this gap by leveraging scRNA-seq references to either resolve spot-level data to single-cell resolution for seq-based ST data or infer transcriptome-wide expression for image-based ST data [73].
In embryonic development, where dynamic gene expression patterns orchestrate cell differentiation and tissue morphogenesis, deconvolution enables researchers to reconstruct spatial gene expression maps at cellular resolution, revealing how transcriptional programs are regulated within the spatial architecture of the embryo. This is particularly crucial for understanding how maternal mRNA gradients establish embryonic axes and how localized translation controls cell fate decisions [74].
Computational Methodologies for Data Integration
Core Mathematical Frameworks
Spatial deconvolution methods employ diverse mathematical approaches to integrate scRNA-seq and ST data. SpatialScope utilizes deep generative models to learn expression distributions of cell types from scRNA-seq reference data, then decomposes spot-level gene expression into single-cell resolution through Langevin dynamics sampling from posterior distributions [73]. The fundamental equation governing this decomposition is:
[ {{{{{{{{\bf{X}}}}}}}}^{(t+1)}={{{{{{{{\bf{X}}}}}}}}}^{(t)}+\eta {\nabla }{{{{{{{{\bf{X}}}}}}}}}\log p\left({{{{{{{{\bf{X}}}}}}}}}^{(t)}| {{{{{{{\bf{y}}}}}}}},\, {k}{1},\, {k}_{2}\right)+\sqrt{2\eta }{{{{{{{{\boldsymbol{\varepsilon }}}}}}}}}^{(t)} ]
where ({{{{{{{\bf{X}}}}}}}}) represents the single-cell expressions, ({{{{{{{\bf{y}}}}}}}}) is the observed spot-level expression, η is the step size, and ε is random noise [73]. This approach allows the decomposition of spot-level gene expression into constituent single-cell expressions when cell types within the spot are known.
The CMAP (Cellular Mapping of Attributes with Position) method employs a divide-and-conquer strategy with three sequential mapping processes: DomainDivision partitions cells into spatial domains using a hidden Markov random field (HMRF) and classifies them with support vector machines; OptimalSpot aligns cells to optimal spots through iterative refinement using a cost function that incorporates structural similarity index (SSIM); and PreciseLocation determines exact cellular coordinates using a spring steady-state model learned from physical fields [68].
For bulk deconvolution, SQUID (Single-cell RNA Quantity Informed Deconvolution) combines RNA-seq transformation and dampened weighted least-squares deconvolution approaches, outperforming ordinary least squares regression, particularly when accurate expression profile estimates are available [75]. ReDeconv addresses critical normalization challenges by incorporating transcriptome size variation across cell types through its Count based on Linearized Transcriptome Size (CLTS) normalization, mitigating scaling effects that distort deconvolution accuracy, especially for rare cell types [76].
Comparative Analysis of Deconvolution Tools
Table 1: Comparison of Key Spatial Deconvolution Methods
| Method | Primary Approach | Spatial Data Compatibility | Key Innovations | Limitations |
|---|---|---|---|---|
| SpatialScope [73] | Deep generative models | seq-based (Visium, Slide-seq) and image-based (MERFISH) | Transcriptome-wide single-cell resolution; Langevin dynamics for decomposition | Computational intensity; Complex implementation |
| CMAP [68] | Divide-and-conquer with HMRF & SVM | Diverse platforms (Xenium, Visium, Slide-seq, seqFISH) | Three-level mapping precision; Spring steady-state model for exact coordinates | Limited benchmarking on embryonic datasets |
| SQUID [75] | Dampened weighted least-squares | Bulk RNA-seq with scRNA-seq reference | RNA quantity informed; Outperforms OLS regression | Designed primarily for bulk deconvolution |
| ReDeconv [76] | Transcriptome size correction | Bulk RNA-seq with scRNA-seq reference | CLTS normalization; Addresses gene length effects | Not specifically designed for spatial data |
| Cell2location [68] | Bayesian modeling | seq-based ST platforms | Cell-type proportion estimation | Limited to spot-level resolution |
| CARD [68] | Reference-based deconvolution | seq-based ST platforms | Spatial domain identification | Requires high-quality reference |
Table 2: Performance Metrics of Deconvolution Methods on Benchmark Datasets
| Method | Cell Usage Ratio | Mapping Accuracy | Rare Cell Detection | Computational Efficiency |
|---|---|---|---|---|
| CMAP [68] | 99% (2215/2242 cells) | 73% weighted accuracy | Good (structural similarity) | Moderate (three-tiered process) |
| CellTrek [68] | 45% (999/2242 cells) | Lower than CMAP | Moderate | High |
| CytoSPACE [68] | 52% (1164/2242 cells) | Lower than CMAP | Moderate | High |
| SpatialScope [73] | Not specified | High (enables ligand-receptor analysis) | Good (generative modeling) | Low (deep learning) |
| SQUID [75] | Not applicable | Superior for bulk deconvolution | Excellent for rare subclones | High |
Experimental Protocols for Method Validation
Cell Mixture Preparation and Characterization
A critical validation approach for deconvolution methods involves preparing in vitro cell mixtures with known compositions to establish ground truth datasets. The following protocol, adapted from systematic evaluation studies, provides a framework for benchmarking deconvolution accuracy [75]:
Cell Line Selection: Select diverse cell types relevant to the research context. For embryonic development, consider including stem cells and differentiated progeny. A representative mixture includes 3 breast cancer lines (T47D, BT474, MCF7), monocytes (Thp1), lymphocytes (Jurkat), and stem cells (hMSC) in varying proportions [75].
Mixture Assembly: Create multiple mixtures with varying proportions of each cell type. Include some mixtures dominated by one cell type (e.g., 66% T47D cells) and others with balanced composition. Maintain consistent percentages of certain types (e.g., 15% monocytes and lymphocytes) while varying others (e.g., 0.5-2% hMSC) to test rare population detection [75].
Validation Profiling: Profile each mixture using multiple modalities:
- Flow cytometry: Perform in triplicate for independent composition evaluation
- Bulk RNA-seq: Profile each cell line and mixture in triplicate
- scRNA-seq: Profile entire mixtures to identify cell type clusters
Data Integration and Correlation: Verify mixture composition by correlating estimates from cell counting, flow cytometry, and scRNA-seq clustering. High correlations (r = 0.96-0.97) between cell counting and other methods validate the ground truth, while lower correlations between flow cytometry and scRNA-seq (r = 0.92) highlight method-specific errors [75].
This approach establishes datasets where expression profiles and composition of each cell type are known with high accuracy, enabling quantitative evaluation of deconvolution methods.
Embryonic Tissue Processing for Spatial Deconvolution
For embryonic studies specifically, special considerations apply for tissue processing and data generation:
Precise Staging: Collect embryos at precisely defined developmental stages. For C. elegans embryos, this might involve synchronization and staging at specific cell cycles [74].
Spatial Transcriptomics Profiling: Apply appropriate spatial technologies based on resolution needs and transcriptome coverage requirements. For comprehensive transcriptome coverage, use seq-based approaches (Visium, Slide-seq); for higher spatial resolution with targeted genes, use image-based approaches (MERFISH, seqFISH) [73].
Reference scRNA-seq Generation: Dissociate replicate embryos into single cells and perform scRNA-seq using 10X Genomics Chromium or Smart-seq2 platforms to create a comprehensive reference atlas [68].
Integration and Validation: Apply deconvolution methods such as SpatialScope or CMAP to integrate scRNA-seq references with spatial data. Validate results using known spatial expression patterns of key developmental genes [73].
The Scientist's Toolkit: Research Reagents and Platforms
Essential Research Reagents and Technologies
Table 3: Essential Research Reagents and Platforms for Spatial Deconvolution
| Category | Specific Technology/Reagent | Function | Application in Embryonic Research |
|---|---|---|---|
| Spatial Transcriptomics Platforms | 10x Visium [73] | Seq-based spatial transcriptomics | Broad transcriptome coverage of embryonic sections |
| Slide-seq [8] | Seq-based high-resolution spatial mapping | Enhanced spatial resolution for embryonic patterning studies | |
| MERFISH [73] | Image-based multiplexed FISH | Targeted high-resolution spatial mapping of key developmental genes | |
| seqFISH [68] | Sequential fluorescence in situ hybridization | Spatial validation of deconvolution results | |
| Single-cell Technologies | 10X Genomics Chromium [68] | High-throughput scRNA-seq | Generating comprehensive reference atlases |
| Smart-seq2 [68] | High-sensitivity full-length scRNA-seq | Detailed characterization of rare cell populations | |
| Computational Tools | SpatialScope [73] | Deep generative model integration | Achieving single-cell resolution from spot-based embryonic data |
| CMAP [68] | Precise cellular mapping | Determining exact spatial coordinates of embryonic cells | |
| SQUID [75] | Bulk deconvolution | Analyzing bulk RNA-seq from whole embryos | |
| ReDeconv [76] | Transcriptome-size-aware deconvolution | Accounting for biological variation in transcriptome size | |
| Specialized Reagents | DNA-barcoded beads [8] | Spatial capture of mRNA | Slide-seq implementation for embryonic tissues |
| Transmitter/receiver beads [8] | Computational array reconstruction | Scalable spatial mapping without imaging |
Experimental Workflow Integration
Applications in Embryonic mRNA Spatial Expression Research
The integration of scRNA-seq data for spatial deconvolution offers particularly powerful applications in embryonic research, where spatial organization is fundamental to developmental processes. Recent studies have demonstrated how regulated translation of maternally deposited mRNAs drives early development, with stage-specific translation patterns emerging despite minimal changes in maternal mRNA abundance [74]. Deconvolution methods enable researchers to spatially resolve these translational regulatory patterns within the embryo architecture.
In practice, SpatialScope has been applied to enhance the resolution of spatial transcriptomics data from human embryonic hematopoietic organoids, producing single-cell resolution data that enabled detection of spatially resolved cell-cell interactions and co-localization of different cell types that were not apparent at spot-level resolution [73]. This approach revealed ligand and receptor pairs essential in vascular proliferation and differentiation in human heart development [73].
For investigating translational regulation in early embryogenesis, methods like SQUID and ReDeconv can be applied to bulk RNA-seq data from staged embryos to deconvolute cell type abundances and dynamics [75] [76]. This is particularly valuable for identifying rare cell populations emerging during differentiation, such as primordial germ cells or specific progenitor populations, whose spatial positioning often determines their developmental fate.
The CMAP approach, with its precise coordinate mapping, enables reconstruction of spatial gene expression gradients that pattern embryonic axes and tissue layers [68]. This capability provides unprecedented insight into how mRNA localization correlates with translational efficiency and how RNA-binding proteins like OMA-1 in C. elegans repress translation of target mRNAs in a stage-specific manner while indirectly promoting translational efficiency of other transcripts [74].
As spatial technologies continue to evolve, the integration of scRNA-seq data for deconvolution will play an increasingly critical role in unraveling the complex spatial regulation of gene expression that orchestrates embryonic development, offering new insights into fundamental biological processes and congenital disorders.
The integration of spatial molecular data with traditional histology represents a paradigm shift in biological research, particularly for understanding complex processes like embryonic development. Hematoxylin and Eosin (H&E) staining has served as the histological cornerstone for over a century, providing fundamental information about tissue architecture and cellular morphology through its differential staining of nuclei (blue/purple) and cytoplasm/extracellular matrix (pink). However, this technique offers limited molecular specificity. In contrast, multiplex immunofluorescence (mIF) enables precise visualization and quantification of multiple protein targets within their native tissue context by using fluorescently-labeled antibodies, while spatial transcriptomics (ST) maps the expression of hundreds to thousands of genes while preserving spatial coordinates [77]. The correlation of these complementary datasets—morphological, proteomic, and transcriptomic—creates a powerful framework for investigating the spatial heterogeneity of tissues, with particular relevance for embryonic research where precise spatiotemporal gene expression dictates organogenesis and tissue patterning [14].
The challenge of detecting mRNA spatial expression in embryos requires sophisticated methodological approaches that preserve spatial context while delivering molecular resolution. Recent advances in spatial omics technologies now enable researchers to create comprehensive "digital embryos" that reconstruct developmental processes at single-cell resolution across three-dimensional space [14]. This technical guide details the methodologies for correlating H&E histology with immunofluorescence and spatial transcriptomic data, providing experimental protocols, analytical frameworks, and practical considerations specifically contextualized for embryonic development research.
Core Technologies and Their Integration
H&E Staining and Multiplex Immunofluorescence
The foundation of spatial correlation begins with high-quality histological preparation. H&E staining remains the universal standard in pathology and research, offering broad morphological context but limited molecular information. Multiplex immunofluorescence expands this capability by enabling simultaneous detection of multiple protein markers within a single tissue section, typically using fluorophore-conjugated antibodies and specialized imaging systems [78]. Advanced platforms like the Orion scanner facilitate this integration by performing whole-slide, 16- to 18-channel IF imaging followed by H&E staining and imaging of the same cells, enabling direct one-to-one comparison of cellular morphology and protein expression patterns [78]. This approach preserves the critical relationship between tissue structure and molecular composition that is essential for understanding embryonic development.
For embryonic research specifically, the combination of multicolor immunofluorescence staining with spatial in situ mRNA expression analysis (e.g., RNAscope) has proven particularly valuable [79]. This methodology allows researchers to simultaneously visualize protein localization via immunofluorescence and mRNA expression patterns via in situ hybridization within the same tissue section, providing direct correlation between transcriptional and translational events in developing tissues.
Spatial Transcriptomics Platforms
Spatial transcriptomics technologies have evolved rapidly, offering increasingly higher resolution for mapping gene expression patterns. Two primary categories dominate the field:
- Sequencing-based approaches (e.g., 10x Visium) utilize spatially barcoded capture probes printed on glass slides to link RNA molecules to their tissue coordinates, followed by next-generation sequencing [80] [77]. These methods provide broad transcriptome coverage but have traditionally been limited to multi-cell resolution (55μm spots with Visium), though newer iterations like Visium HD are improving resolution.
- Imaging-based approaches (e.g., 10x Xenium, NanoString CosMx, Vizgen MERSCOPE) use in situ hybridization to visualize hundreds to thousands of RNA species directly in tissue sections, achieving subcellular resolution [81] [77]. These methods offer superior spatial resolution but typically profile a smaller subset of the transcriptome.
For embryonic research, the choice between platforms depends on the specific research question, balancing the need for whole-transcriptome coverage against spatial resolution requirements. Recent work on mouse organogenesis has demonstrated the power of combining these approaches with histology to reconstruct full digital embryos at single-cell resolution during critical developmental windows [14].
Table 1: Comparison of Primary Spatial Technologies for Embryonic Research
| Technology | Resolution | Molecular Targets | Throughput | Best Applications in Embryonic Research |
|---|---|---|---|---|
| H&E Staining | Subcellular | Tissue morphology | High | Initial histological assessment, tissue architecture reference |
| Multiplex IF | Subcellular | 4-100+ proteins | Medium | Protein co-localization, cell typing, signaling pathway analysis |
| 10x Visium | 55μm spots (multi-cell) | Whole transcriptome | High | Exploratory gene expression patterning in developing tissues |
| 10x Xenium | Subcellular | 100s-1000s of genes | Medium | High-resolution mapping of specific gene panels at cellular level |
| RNAscope | Single RNA molecules | 1-12 RNA targets | Medium | Validation of key developmental genes, low-abundance transcripts |
Integrated Experimental Workflows
Correlative H&E and Multiplex Immunofluorescence
The workflow for correlating H&E morphology with multiplex immunofluorescence begins with careful tissue preparation. For embryonic tissues, optimal fixation conditions must balance preservation of morphology with antigen accessibility. Following standard H&E staining and imaging, tissues can be destained and subjected to multiplex immunofluorescence protocols. Advanced approaches like the Orion method enable sequential acquisition of high-plex IF images followed by H&E staining on the same tissue section, ensuring perfect registration between molecular and morphological data [78].
For embryonic applications, panel design should prioritize markers relevant to developmental processes, such as lineage-specific transcription factors, signaling molecules, and differentiation markers. A typical workflow might include:
- Tissue preparation and sectioning of embryonic specimens at defined developmental stages
- H&E staining and high-resolution imaging to establish morphological reference
- Multiplex immunofluorescence using antibodies against key developmental markers
- Image registration and analysis to correlate protein expression with tissue architecture
Recent methodological innovations like SEPARATE (Spatial Expression PAttern-guided paiRing And unmixing of proTEins) can further enhance multiplexing capability by pairing proteins with distinct spatial expression patterns, allowing two proteins to be imaged using a single fluorophore and subsequently computationally unmixed based on their spatial patterns [82]. This approach effectively doubles the multiplexing capacity without additional staining cycles, particularly valuable for precious embryonic specimens.
Figure 1: Integrated workflow for correlative H&E, multiplex immunofluorescence, and spatial transcriptomics analysis of embryonic tissues.
Spatial Transcriptomics with Histological Integration
For spatial transcriptomics, the workflow varies depending on the platform but shares common elements. The 10x Visium platform, for example, involves mounting tissue sections onto spatially barcoded slides, performing H&E staining and imaging, followed by tissue permeabilization to release RNA which is captured by the barcoded oligos, and finally library preparation and sequencing [80]. The resulting gene expression data is inherently aligned with the H&E image through the spatial barcoding system.
For embryonic research, studies have demonstrated the power of combining serial sectioning with ST profiling to reconstruct three-dimensional transcriptional landscapes. In groundbreaking work on mouse organogenesis, researchers profiled 285 serial sections from E7.5–E8.0 embryos to generate full spatiotemporal transcriptome maps during early organogenesis at single-cell resolution [14]. This approach, combined with computational reconstruction methods like SEU-3D, enables the creation of comprehensive "digital embryos" that preserve spatial context while providing whole-transcriptome data.
The integration of H&E histology with spatial transcriptomics is particularly powerful for identifying specialized microenvironments or niches within developing embryos. In endometrial research, for example, ST analysis integrated with H&E morphology identified seven distinct cellular niches with specific gene expression characteristics [80]. Similar approaches can be applied to embryonic tissues to understand the molecular signatures of emerging organ primordia and signaling centers.
Computational Integration and Analysis
Data Registration and Multi-modal Correlation
The correlation of H&E, immunofluorescence, and spatial transcriptomics data requires robust computational methods for image registration and data integration. When using platforms like Orion that generate H&E and mIF from the same tissue section, registration is inherent to the methodology [78]. However, when data originates from serial sections or different technologies, computational alignment becomes essential.
For embryonic research, where structures may be small and rapidly changing, landmark-based registration approaches can align H&E and IF images based on distinctive morphological features. More advanced methods use deep learning-based registration to achieve precise alignment even when tissue deformation occurs between sections. Once registered, the correlated data enables quantitative analysis of relationships between tissue morphology, protein expression, and gene expression patterns.
Spatial analysis tools originally developed for ecology and geography have been adapted for biological applications, providing robust methods for quantifying spatial patterns in multiplexed tissue data [83]. The pair correlation function, for example, can identify clustered, random, or dispersed distributions of specific cell types within embryonic tissues, while the nearest neighbor G-function examines the cumulative distribution of distances between cells of the same or different phenotypes.
Predicting Spatial Gene Expression from Histology
A particularly promising area of computational integration is the prediction of spatial gene expression patterns directly from H&E histology images. Deep learning methods like GHIST (spatial gene expression from HISTology) leverage the relationship between tissue morphology and gene expression to predict spatially-resolved single-cell gene expression from routine H&E images [81]. This approach uses a multitask architecture that considers interdependencies between cell type, neighborhood composition, nucleus morphology, and single-cell RNA expression.
For embryonic research, where spatial transcriptomics experiments may be cost-prohibitive at large scales, such methods enable researchers to leverage existing H&E archives to generate hypotheses about spatial gene expression patterns. Validation studies have shown that GHIST effectively captures single-cell spatial expression, with high correlation between predicted and measured expression for spatially variable genes (median correlation of 0.7 for top genes) [81].
Similarly, methods like MIPHEI (Multiplex Immunofluorescence Prediction from H&E) use Vision Transformer foundation models in a U-Net architecture to predict protein expression patterns from H&E morphology alone [84]. This approach has demonstrated accurate cell-type classification from H&E alone, with F1 scores of 0.88 for Pan-CK and 0.57 for CD3e, substantially outperforming baseline methods for most markers.
Figure 2: Computational workflow for predicting spatial molecular data from H&E histology using multi-task deep learning architectures like GHIST and MIPHEI.
Application to Embryonic mRNA Spatial Expression
Case Study: Digital Reconstruction of Mouse Embryogenesis
A landmark application of correlative spatial methods in embryonic research comes from the digital reconstruction of full mouse embryos during early organogenesis (E7.5–E8.0) [14]. This study employed serial spatial transcriptomics profiling of 285 sections from six embryos, integrated with histological analysis to create comprehensive spatiotemporal atlases of mesoderm and endoderm lineages. The researchers developed SEU-3D, a computational method for reconstructing three-dimensional digital embryos from two-dimensional sections, enabling investigation of regionalized gene expression in the native spatial context.
This integrated approach revealed previously uncharacterized aspects of early organogenesis, including the identification of a primordium determination zone (PDZ) at the embryonic-extraembryonic interface at E7.75 [14]. Through correlation of spatial gene expression patterns with tissue morphology, the study demonstrated that coordinated signaling communications across germ layers contribute to the formation of cardiac primordium. This work exemplifies the power of combining spatial molecular profiling with histological context to uncover new biological principles in embryonic development.
Technical Considerations for Embryonic Tissues
Working with embryonic tissues presents unique challenges for spatial technologies. Embryonic specimens are often small, delicate, and exhibit rapid changes in gene expression and morphology. Key considerations include:
- Optimal fixation conditions that preserve both morphology and RNA/protein integrity without excessive crosslinking
- Section thickness optimization to balance morphological preservation with molecular accessibility
- Stage-matched sampling to account for rapid developmental transitions
- Three-dimensional reconstruction from serial sections to understand spatial relationships in the context of embryonic structure
For mRNA detection specifically, methods like RNAscope provide high sensitivity and single-molecule resolution for key developmental genes [85]. The proprietary "double Z" probe design enables highly specific and sensitive detection of target RNA, with each dot representing a single RNA transcript. This approach can be combined with immunofluorescence to correlate protein and mRNA localization within the same embryonic tissue section.
Table 2: Research Reagent Solutions for Embryonic Spatial Analysis
| Reagent Category | Specific Examples | Function in Embryonic Research | Technical Considerations |
|---|---|---|---|
| Antibody Panels | Transcription factors (Oct4, Nanog), Signaling molecules (BMP4, Wnt), Lineage markers (Sox17, T brachyury) | Cell type identification, lineage tracing, signaling pathway activity | Validate specificity in embryonic tissues; consider species cross-reactivity |
| Spatial Transcriptomics Kits | 10x Visium Spatial Gene Expression, 10x Xenium Gene Panels | Whole transcriptome mapping, targeted gene panel analysis | Optimize permeabilization for embryonic tissues; select species-specific probes |
| In Situ Hybridization Probes | RNAscope probes, Molecular Instruments programmable probes | mRNA localization with single-molecule sensitivity | Design against specific splice variants; confirm sequence specificity |
| Multiplexing Reagents | Akoya PhenoCycler antibodies, SEPARATE pairing reagents | Expanding multiplexing capacity for protein detection | Validate fluorophore compatibility; optimize staining cycles for embryonic tissues |
| Image Registration Tools | Orion platform, ASHLAR, BigStitcher | Multi-modal image alignment and correlation | Account for tissue deformation between sections; use fiduciary markers when possible |
The correlation of spatial data with histology represents a transformative approach for investigating mRNA spatial expression in embryonic development. As technologies continue to advance, we anticipate several key developments that will further enhance this field:
First, the integration of multiple molecular modalities—transcriptome, proteome, epigenome—within the same embryonic tissue section will provide increasingly comprehensive views of developmental regulation. Methods like SEPARATE that enable expanded multiplexing through computational unmixing [82] will play a crucial role in this integration.
Second, computational prediction methods like GHIST [81] and MIPHEI [84] will mature, enabling researchers to extract increasingly accurate spatial molecular information from routine H&E histology. This will be particularly valuable for leveraging historical collections of embryonic tissue sections for spatial analysis.
Third, three-dimensional reconstruction methodologies will improve, allowing true volumetric analysis of gene expression patterns throughout developing embryos. The digital embryo reconstruction pioneered in mouse studies [14] will extend to other model organisms and human embryonic tissues where available.
In conclusion, the correlation of H&E staining and immunofluorescence with spatial transcriptomics provides a powerful multi-modal framework for understanding the spatial regulation of gene expression during embryonic development. By integrating morphological context with molecular precision, these approaches are revealing the complex spatiotemporal dynamics that guide organogenesis and tissue patterning. As methods continue to evolve, they will undoubtedly yield new insights into the fundamental principles of development and the spatial organization of biological systems.
The emergence of spatial transcriptomics has revolutionized our understanding of gene expression by allowing researchers to profile the entire transcriptome while preserving the spatial context of cells within tissues. However, this technological advancement brings forth a critical challenge: how to validate, benchmark, and compare results across different platforms, experiments, and laboratories. The establishment of gold standards through well-characterized reference tissues and defined anatomical regions provides the essential solution to this challenge, creating a common framework that ensures data reliability, reproducibility, and interoperability. Within embryo research, where spatial gene expression patterns dictate developmental fate, these reference standards become particularly crucial for distinguishing technical artifacts from biologically significant findings, enabling the scientific community to build upon a foundation of verified spatial mRNA expression data.
The Critical Need for Reference Standards in Spatial Biology
Spatial transcriptomics technologies have rapidly diversified, with at least 11 distinct sequencing-based methods currently available, each with varying capabilities in spatial resolution, capture efficiency, and molecular diffusion rates [62]. This methodological plurality, while driving innovation, creates substantial challenges in cross-platform validation and data integration. Without standardized references, findings from one technological platform remain siloed and difficult to compare with results from another platform.
The problem extends beyond mere technical compatibility. Biological interpretation of spatial transcriptomic data requires accurate annotation of spatial domains and cell types, tasks that are highly dependent on the quality and resolution of the underlying data [7] [62]. Reference tissues with well-defined anatomical structures provide the ground truth against which computational tools for spatial analysis can be benchmarked, ensuring that identified spatial expression patterns reflect biology rather than analytical artifacts.
For embryonic development research, where mRNA localization often precedes and determines protein expression and cellular fate, the ability to reliably track these patterns across developmental timepoints is paramount [32] [86]. The establishment of standardized reference tissues enables longitudinal studies of mRNA expression dynamics and enables collaboration across research groups studying similar biological questions.
Characteristics of Ideal Reference Tissues
Ideal reference tissues for spatial transcriptomics possess specific characteristics that make them suitable for standardization purposes. These features ensure that the tissues can serve as reliable benchmarks across platforms and experiments.
Well-Defined Anatomy and Histology
Tissues with clearly delineated anatomical substructures that are consistent across individuals allow for precise orientation and mapping of spatial data. For example, the mouse hippocampus exhibits consistent thickness and contains distinct subregions including CA1, CA2, CA3, and the dentate gyrus, each with known cellular composition and gene expression profiles [62]. Similarly, E12.5 mouse eyes in embryo present a known structure with a lens surrounded by neuronal retina cells, providing clearly identifiable landmarks for spatial mapping.
Conserved Expression Patterns
Reference tissues should contain regions with established, conserved gene expression patterns that can serve as positive controls for spatial technologies. The mouse olfactory bulb features clear layer separation with various neuron types expressing known marker genes, allowing researchers to verify that their platforms can detect expected spatial expression gradients [62].
Structural Stability Across Development
For embryonic studies, tissues that maintain core architectural features across developmental timepoints enable the tracking of expression dynamics within a consistent spatial framework. The zebrafish one-cell embryo, with its clearly established animal-vegetal axis, provides a model system for studying the initial asymmetries that establish embryonic patterning [32].
Practical Handling Properties
From a practical standpoint, ideal reference tissues should be readily obtainable, easily sectioned, and maintain structural integrity through processing steps. Mouse brain tissues, for instance, are widely used because they are relatively homogeneous across individuals and can be consistently sectioned to reveal the same anatomical landmarks [62].
Established Reference Tissues in Current Research
Several tissues have emerged as standard references in spatial transcriptomics research, each offering unique advantages for specific applications. The table below summarizes key reference tissues and their applications in spatial transcriptomics studies.
Table 1: Established Reference Tissues for Spatial Transcriptomics
| Tissue Type | Defined Anatomical Regions | Key Marker Genes | Applications in Validation |
|---|---|---|---|
| Mouse Brain | Hippocampus (CA1, CA2, CA3, dentate gyrus), cortical layers, thalamic nuclei | Prdm8, Prox1, Slc17a7 [62] | Platform sensitivity, spatial resolution, cell type mapping |
| Mouse Olfactory Bulb | Distinct laminar organization with various neuron types | Layer-specific markers | Resolution limits, domain identification |
| E12.5 Mouse Embryo | Developing eye (lens, neuronal retina), limb buds, somites | Vit, Crybb3 (lens), Aldh1a1 (neural retina) [62] | Morphological patterning, developmental gene expression |
| Zebrafish One-Cell Embryo | Animal-vegetal axis, blastodisc | dazl, trim36, grip2a, wnt8a [32] | mRNA localization, embryonic polarization |
| Drosophila Ovary | Oocyte, nurse cells, somatic epithelial cells | oskar, bicoid, gurken [87] | Subcellular mRNA transport, translational control |
Experimental Protocols for Reference Tissue Analysis
Standardized protocols for reference tissue preparation and analysis are essential for generating comparable data across laboratories and platforms. The following sections detail established methodologies for different tissue types and applications.
Spatial Transcriptomics of FFPE Reference Tissues
Formalin-fixed paraffin-embedded (FFPE) tissues represent the vast majority of clinical biospecimens in biobanks, making them invaluable for retrospective studies. A validated protocol for spatial transcriptomics of FFPE tissues includes the following key steps [56]:
Tissue Sectioning and Attachment: Float FFPE tissue sections onto spatially barcoded slides, then dry in an oven to ensure adhesion.
Deparaffinization: Immerse slides successively in xylene and ethanol to remove paraffin while preserving tissue morphology.
Histological Staining: Stain with hematoxylin and eosin (H&E) and image under a high-resolution microscope for morphological reference.
Cross-link Reversal: Perform heat-induced retrieval at 70°C with Tris-EDTA buffer at pH 8.0 to reverse formalin-induced cross-links. A pH of 8.0 prevents unwanted side reactions such as pH-dependent RNA hydrolysis.
Tissue Permeabilization: Treat with collagenase followed by enzymatic permeabilization optimized based on tissue type and determined through tissue optimization assays.
mRNA Capture and Library Preparation: Capture mRNA using spatially barcoded oligo(dT) probes, followed by cDNA synthesis and library preparation for sequencing.
This protocol has been successfully applied to mouse brain sections, ovarian carcinosarcoma samples, human organoids, and SARS-CoV-2-infected lung tissue, demonstrating its versatility across tissue types [56].
Systematic mRNA Localization Mapping in Drosophila Ovaries
For studying mRNA localization in developmental models, the Drosophila ovary provides an excellent system. A comprehensive protocol for genome-wide mapping includes [87]:
Tissue Collection and Fixation: Mass isolate egg chambers from Drosophila ovaries and fix to preserve tissue architecture and mRNA locations.
Stage-Specific mRNA Sequencing: Perform 3Pseq and RNAseq on staged egg chambers to obtain transcriptional profiles.
Fluorescent In Situ Hybridization (FISH): Conduct genome-wide FISH using tagged antisense riboprobes against thousands of mRNAs.
High-Resolution Imaging: Acquire three-dimensional image stacks of hybridized tissues using fluorescence microscopy.
Pattern Annotation and Data Integration: Systematically annotate expression patterns using a hierarchical controlled vocabulary and integrate with sequencing data in a searchable database (Dresden Ovary Table).
This approach has revealed that mRNA localization is widespread in the ovary, with 64% of expressed genes showing ubiquitous distribution, 22% exhibiting subcellular localization, and 13% showing cell-type-specific expression [87].
Tomo-seq for Spatial Mapping in Zebrafish Embryos
The tomoseq method provides high-resolution spatial mapping of gene expression in early embryos through a systematic sectioning approach [32]:
Embryo Orientation and Embedding: Embed and orient individual zebrafish embryos at the one-cell stage along the animal-vegetal axis.
Cryosectioning: Section the embryo and yolk sac into thin slices (96 sections per embryo) using a cryostat.
RNA Extraction and Barcoding: Extract RNA from each section and barcode with unique molecular identifiers.
Library Preparation and Sequencing: Prepare sequencing libraries and sequence to generate spatial expression data.
Data Analysis and Pattern Identification: Map sequencing reads to the genome, assign to spatial bins, and identify localization patterns using self-organizing maps or other clustering approaches.
This method enabled identification of 97 genes specifically localized at the vegetal pole of the one-cell zebrafish embryo, increasing the number of known vegetal genes by approximately tenfold [32].
Visualization of Tissue Selection Criteria
The following diagram illustrates the decision process for selecting appropriate reference tissues based on research goals and tissue characteristics:
Tissue Selection Criteria Flowchart
Analytical Frameworks for Reference Data
The value of reference tissues is fully realized only when paired with robust analytical frameworks that can extract meaningful biological insights from spatial data. Several computational approaches have been developed specifically for this purpose.
Spatial Data Alignment and Integration
Multiple tissue sections from reference tissues require alignment and integration to reconstruct three-dimensional contexts or combine data across experiments. Computational tools for this task fall into three main categories [7]:
Table 2: Computational Approaches for Spatial Data Integration
| Method Category | Representative Tools | Key Applications | Strengths |
|---|---|---|---|
| Statistical Mapping | PASTE, GPSA, Eggplant | Spatial differential expression, 3D mapping | Statistical rigor, handling technical variability |
| Image Processing & Registration | STalign, STIM, STUtility | Cell-type identification, spatial clustering | Leverages histological information |
| Graph-Based Methods | SpatiAlign, STAligner, Graspot | Cellular trajectory, spatial domain identification | Captures spatial neighborhood relationships |
These tools enable researchers to align multiple sections of reference tissues like the mouse brain, creating comprehensive 3D models of gene expression that preserve spatial relationships across the entire tissue architecture [7].
Benchmarking Spatial Technologies
Reference tissues serve as critical substrates for comparing the performance of different spatial transcriptomics platforms. A standardized benchmarking approach involves [62]:
Processing identical reference tissues across multiple platforms (e.g., Visium, Stereo-seq, Slide-seqV2, DBiT-seq)
Measuring key performance parameters including:
- Molecular diffusion rates
- Capture efficiency (sensitivity)
- Spatial resolution
- Gene detection sensitivity
Assessing biological inference capabilities for tasks such as:
- Region annotation accuracy
- Cell-type identification
- Marker gene detection
- Cell-cell communication inference
Using this approach, researchers have demonstrated that spatial transcriptomic data possess unique attributes beyond merely adding a spatial axis to single-cell data, including enhanced ability to capture patterned rare cell states [62].
Essential Research Reagents and Platforms
The experimental workflows for reference tissue analysis depend on specialized reagents and platforms. The following table catalogues key solutions used in spatial transcriptomics studies.
Table 3: Essential Research Reagents and Platforms for Spatial Transcriptomics
| Reagent/Platform | Type | Function | Example Applications |
|---|---|---|---|
| Visium Spatial Gene Expression | Platform | Genome-wide spatial mapping using spatially barcoded oligo arrays | Mouse brain mapping, cancer microenvironment [62] [88] |
| Xenium In Situ | Platform | Subcellular resolution spatial analysis using in situ hybridization | Cellular architecture in mouse neuroinflammation models [88] |
| Tris-EDTA Buffer (pH 8.0) | Reagent | Cross-link reversal in FFPE tissues | mRNA recovery from archived clinical samples [56] |
| Template-Switch Oligo (TSO) | Reagent | cDNA amplification during reverse transcription | Quality control for FFPE spatial transcriptomics [56] |
| 4-thiouridine (4sU) | Reagent | Metabolic RNA labeling for distinguishing maternal/zygotic transcripts | Tracking mRNA fate in zebrafish development [32] |
| Collagenase | Reagent | Tissue permeabilization for enhanced mRNA accessibility | FFPE tissue processing for spatial transcriptomics [56] |
| Antisense Riboprobes | Reagent | Target-specific detection in FISH assays | Genome-scale mRNA localization in Drosophila [87] |
Visualization of Experimental Workflow
The following diagram illustrates a generalized experimental workflow for spatial transcriptomics analysis of reference tissues:
Spatial Transcriptomics Workflow
Reference tissues with defined anatomical regions constitute the cornerstone of rigorous spatial transcriptomics, providing the standardized framework necessary for technology benchmarking, method validation, and biological discovery. As spatial technologies continue to evolve toward higher resolution and greater sensitivity, these biological standards will only grow in importance, ensuring that findings across platforms and laboratories can be integrated into a coherent understanding of spatial gene expression architecture. For embryonic development research, where precise mRNA localization dictates cellular fate and tissue patterning, reference tissues like the zebrafish embryo and Drosophila ovary provide indispensable models for deciphering the spatial code of development. The continued refinement of both the reference tissues themselves and the analytical frameworks for their interpretation will accelerate progress toward a comprehensive spatial atlas of gene expression across development and disease.
Conclusion
Spatial transcriptomics has fundamentally transformed our ability to decipher the complex molecular architecture of the developing embryo, moving beyond single-cell data to provide essential contextual information. The systematic benchmarking of various platforms offers clear guidance for method selection based on specific research goals, balancing resolution, sensitivity, and throughput. As computational tools for data integration and analysis mature, the potential to create comprehensive 'digital embryos' becomes increasingly attainable. Future directions will focus on achieving higher multiplexing, single-cell resolution at transcriptome-wide depth, and dynamic tracking of mRNA fate. These advancements promise to unlock profound insights into congenital disorders, refine drug development processes, and ultimately build a spatially-resolved atlas of human development, marking a new era in developmental biology and regenerative medicine.
References
- 1. What is spatial transcriptomics? [https://sangerinstitute.blog/2025/10/22/what-is-spatial-transcriptomics/]
- 2. Opportunities and challenges in the application of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12568615/]
- 3. A practical guide for choosing an optimal spatial ... [https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-025-11235-3]
- 4. Across the space: applications of spatial transcriptomic ... [https://www.frontiersin.org/journals/cell-and-developmental-biology/articles/10.3389/fcell.2025.1656918/full]
- 5. Spatial Transcriptomics Overview [https://nanostring.com/blog/what-is-spatial-gene-expression/]
- 6. The spatial transcriptome of the late-stage embryonic and ... [https://www.nature.com/articles/s41598-025-95496-8]
- 7. A comprehensive review of spatial transcriptomics data ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12199153/]
- 8. Scientists have developed a way to scale up spatial ... [https://www.broadinstitute.org/news/scaling-up-spatial-genomics]
- 9. Spatial Biology Market Analysis and Forecast, 2025-2035 [https://www.businesswire.com/news/home/20251124442495/en/Spatial-Biology-Market-Analysis-and-Forecast-2025-2035-Opportunities-in-AI-Bridging-the-Gap-Between-Routine-Pathology-and-Spatial-Omics-Technology-and-Partnerships-Boosting-Spatial-Biology---ResearchAndMarkets.com]
- 10. Beyond cell atlases: spatial biology reveals mechanisms ... [https://www.nature.com/articles/s41587-025-02699-5]
- 11. Spatial transcriptomics reveals human cortical layer and ... [https://www.nature.com/articles/s41586-025-09010-1]
- 12. Spatiotemporal transcriptomic maps of whole mouse ... [https://www.nature.com/articles/s41588-023-01435-6]
- 13. Spatial omics advances for in situ RNA biology [https://www.sciencedirect.com/science/article/pii/S1097276524006567]
- 14. Digital reconstruction of full embryos during early mouse ... [https://www.sciencedirect.com/science/article/pii/S0092867425006221]
- 15. Whole‐Gut Spatial Genomic Analysis Reveals Molecular ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12412201/]
- 16. Phenotypically anchored transcriptomics across diverse ... [https://www.frontiersin.org/journals/toxicology/articles/10.3389/ftox.2025.1675060/full]
- 17. Cell-type-specific mRNA transcription and degradation ... [https://www.nature.com/articles/s41467-024-47290-9]
- 18. Sequential and additive expression of miR-9 precursors ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9641661/]
- 19. A computational pipeline for spatial mechano-transcriptomics [https://www.nature.com/articles/s41592-025-02618-1]
- 20. Spatiotemporal dynamics and selectivity of mRNA translation ... [https://pubmed.ncbi.nlm.nih.gov/40985772/]
- 21. CRISPR/Cas-mediated mRNA knockdown in the embryos of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12020200/]
- 22. Searchable Databases of Xenopus RNA-Seq Expression ... [https://www.xenbase.org/xenbase/doNewsRead.do?id=214]
- 23. A new alternative method using cyp3a65 expression in ... [https://www.sciencedirect.com/science/article/pii/S0160412025006233?dgcid=rss_sd_all]
- 24. EMB is essential for enteric nervous system development ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12465749/]
- 25. Tracing the enteric neural crest cell pathway from origin to ... [https://link.springer.com/article/10.1007/s11033-025-11211-w]
- 26. High-resolution spatial transcriptomics and cell lineage ... [https://www.nature.com/articles/s41467-025-59206-2]
- 27. Across the space: applications of spatial transcriptomic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12583089/]
- 28. Integration of single-cell and spatial transcriptomics by SEU ... [https://genomebiology.biomedcentral.com/articles/10.1186/s13059-025-03633-3]
- 29. ELLA: modeling subcellular spatial variation of gene ... [https://www.nature.com/articles/s41467-025-64867-0]
- 30. Visium Spatial Platform | 10x Genomics [https://www.10xgenomics.com/platforms/visium]
- 31. STOmics Stereo-seq [https://en.stomics.tech/why-stereo-seq/index.html]
- 32. Spatio-temporal mRNA tracking in the early zebrafish embryo [https://www.nature.com/articles/s41467-021-23834-1]
- 33. Visium Spatial Assays | 10x Genomics [https://www.10xgenomics.com/platforms/visium/product-family]
- 34. High-definition spatial transcriptomic profiling of immune ... [https://www.nature.com/articles/s41588-025-02193-3]
- 35. STOmics Stereo-seq | Advanced Spatial Multi-omics Technology [https://en.stomics.tech/]
- 36. Analysis, visualization, and integration of spatial datasets ... [https://satijalab.org/seurat/articles/spatial_vignette.html]
- 37. Spaco: A comprehensive tool for coloring spatial data at ... [https://www.sciencedirect.com/science/article/pii/S2666389923003240]
- 38. Visualizing Spatial Transcriptomics: A Guide to Effective Plotting [https://nanostring-biostats.github.io/CosMx-Analysis-Scratch-Space/posts/spatial-plotting/]
- 39. An introduction to spatial transcriptomics for biomedical research [https://genomemedicine.biomedcentral.com/articles/10.1186/s13073-022-01075-1]
- 40. Concordance of MERFISH with bulk and... spatial transcriptomics [https://pmc.ncbi.nlm.nih.gov/articles/PMC9760489/]
- 41. seqFISH - Cai Lab [https://spatial.caltech.edu/seqfish/]
- 42. Emerging Technologies in Tissue Gene Expression Profiling [https://pmc.ncbi.nlm.nih.gov/articles/PMC10603609/]
- 43. What is Spatial Transcriptomics for Spatial Biology? KeyInfo [https://www.completegenomics.com/what-is-spatial-transcriptomics/]
- 44. | Vizgen MERFISH Spatial Transcriptomics [https://vizgen.com/technology/]
- 45. Probes for MERFISH - Synoligo Spatial Transcriptomics [https://synoligo.com/merfish-probes-for-spatial-transcriptomics/]
- 46. Technology | Spatial Genomics | Decoding molecular identity [https://spatialgenomics.com/technology/]
- 47. Spatial transcriptomics levels up | Nature Methods [https://www.nature.com/articles/s41592-019-0441-8]
- 48. - Wikipedia Spatial transcriptomics [https://en.wikipedia.org/wiki/Spatial_transcriptomics]
- 49. Sequencing vs. Imaging in Spatial Transcriptomics: How to Choose the... [https://www.scdiscoveries.com/blog/sequencing-vs-imaging-in-spatial-transcriptomics/]
- 50. Thor: a platform for cell-level investigation of spatial ... transcriptomics [https://www.nature.com/articles/s41467-025-62593-1?error=cookies_not_supported&code=8d34742b-6059-4d88-aae7-1a17b00bf0f9]
- 51. Predictive and robust gene for selection transcriptomics spatial [https://www.nature.com/articles/s41467-023-37392-1?error=cookies_not_supported&code=45f7375d-4ec7-4c80-b864-3431ca823e2a]
- 52. Advances in spatial transcriptomics and its application ... [https://www.nature.com/articles/s41413-025-00429-w]
- 53. A spatiotemporal atlas of mouse gastrulation and early ... [https://www.sciencedirect.com/science/article/pii/S2211124725008186]
- 54. Smad4 controls signaling robustness and morphogenesis ... [https://www.nature.com/articles/s41467-021-26486-3]
- 55. Signaling Pathways in Human Blastocyst Development [https://pmc.ncbi.nlm.nih.gov/articles/PMC12452331/]
- 56. Genome-wide spatial expression profiling in formalin-fixed ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9903805/]
- 57. Enhancing RNA Capture Efficiency in Spatial Transcriptomics [https://pmc.ncbi.nlm.nih.gov/articles/PMC12652657/]
- 58. Mapping human tissues with highly multiplexed RNA in situ ... [https://www.nature.com/articles/s41467-024-46437-y]
- 59. Protocol for single-cell spatial transcriptomic profiling of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12491524/]
- 60. visualizing and understanding mRNA localization [https://pmc.ncbi.nlm.nih.gov/articles/PMC4484810/]
- 61. Frontiers Publishing Partnerships | Advancements in single-cell RNA ... [https://www.frontierspartnerships.org/journals/acta-biochimica-polonica/articles/10.3389/abp.2025.13922/full]
- 62. Systematic comparison of sequencing-based spatial ... [https://www.nature.com/articles/s41592-024-02325-3]
- 63. Budgeting for an mRNA -seq project? How much does RNA Seq cost ? [https://alitheagenomics.com/blog/budgeting-for-an-mrna-seq-project-here-are-the-main-cost-drivers-to-keep-an-eye-on]
- 64. Multidimensional quantitative analysis of mRNA expression ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5825878/]
- 65. DIVERSE SPATIAL EXPRESSION PATTERNS EMERGE ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6779125/]
- 66. Deep learning-based models for preimplantation mouse ... [https://www.nature.com/articles/s41592-024-02511-3]
- 67. From pixels to cell types - PubMed Central - NIH [https://pmc.ncbi.nlm.nih.gov/articles/PMC12577344/]
- 68. High-resolution mapping of single cells in spatial context [https://www.nature.com/articles/s41467-025-61667-4]
- 69. Systematic benchmarking of high-throughput subcellular ... [https://www.nature.com/articles/s41467-025-64292-3]
- 70. Systematic benchmarking of imaging spatial ... [https://www.nature.com/articles/s41467-025-64990-y]
- 71. Driving Innovation in Spatial Transcriptomics [https://bonsailab.com/driving-innovation-in-spatial-transcriptomics-benchmarking-high-throughput-platforms/]
- 72. Comparison of imaging based single-cell resolution spatial ... [https://www.nature.com/articles/s41467-025-63414-1]
- 73. Integrating spatial and single-cell transcriptomics data ... [https://www.nature.com/articles/s41467-023-43629-w]
- 74. Landscape and regulation of mRNA translation in the early ... [https://www.sciencedirect.com/science/article/pii/S2211124725005492]
- 75. Effective methods for bulk RNA-seq deconvolution using ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10394903/]
- 76. Transcriptome size matters for single-cell RNA-seq ... [https://www.nature.com/articles/s41467-025-56623-1]
- 77. The Rise of Spatial Omics in Immunology - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC12501406/]
- 78. High-plex immunofluorescence imaging and traditional ... [https://www.nature.com/articles/s43018-023-00576-1]
- 79. Combined multi-color immunofluorescence staining and ... [https://www.sciencedirect.com/science/article/pii/S0023683725001515]
- 80. A spatial transcriptomics dataset of the endometrium from ... [https://www.nature.com/articles/s41597-025-05995-6]
- 81. Spatial gene expression at single-cell resolution from ... [https://www.nature.com/articles/s41592-025-02795-z]
- 82. Doubling multiplexed imaging capability via spatial ... [https://www.nature.com/articles/s42003-025-08357-5]
- 83. Spatial modelling of the tumor microenvironment from ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10765501/]
- 84. MIPHEI-vit: Multiplex Immunofluorescence Prediction from ... [https://arxiv.org/html/2505.10294v1]
- 85. 2025 Jan 14 - Unlock the role of spatial profiling in ... [https://acdbio.com/2025-jan-14-unlock-role-spatial-profiling-detecting-diverse-mrna-markers-and-short-targets]
- 86. Principles and roles of mRNA localization in animal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3424039/]
- 87. Systematic imaging reveals features and changing ... [https://elifesciences.org/articles/05003]
- 88. Analysis Approaches for Spatial Gene Expression Data [https://www.10xgenomics.com/analysis-guides/spatial-gex-lit-review]