Unlocking Single-Molecule Detection: How RNAscope's Sensitivity Surpasses Traditional ISH
This article provides a comprehensive analysis for researchers and drug development professionals on the significant advancements in in situ hybridization (ISH) sensitivity offered by RNAscope technology.
Unlocking Single-Molecule Detection: How RNAscope's Sensitivity Surpasses Traditional ISH
Abstract
This article provides a comprehensive analysis for researchers and drug development professionals on the significant advancements in in situ hybridization (ISH) sensitivity offered by RNAscope technology. We explore the foundational principles behind its proprietary 'double Z' probe design that enables single-molecule RNA visualization, a capability largely unattainable with traditional ISH. The content details practical methodologies and diverse applications across oncology, neuroscience, and infectious disease research, supported by validation studies demonstrating high concordance with PCR and NGS. Furthermore, we address troubleshooting for assay optimization and discuss the transformative implications of this enhanced sensitivity for biomarker discovery, therapeutic development, and clinical diagnostics.
The Sensitivity Gap: Why Traditional ISH Falls Short and How RNAscope Bridges It
For decades, traditional in situ hybridization (ISH) techniques have been plagued by significant limitations that hindered their reliability for sensitive RNA detection in research and clinical settings. The fundamental challenge stems from the inherent instability of RNA molecules, which are susceptible to degradation by ubiquitous RNases, coupled with the technical constraints of legacy ISH methods [1]. These methods often suffered from poor signal-to-noise ratios due to non-specific probe binding, leading to insufficient sensitivity and specificity for detecting many biologically important RNA targets, especially those with low expression levels [2] [3]. This technological gap created a critical need for a more robust spatial biology platform that could deliver single-molecule sensitivity while preserving tissue morphology and cellular context.
The introduction of RNAscope technology (Advanced Cell Diagnostics) represented a paradigm shift in spatial genomics. Its novel design directly addresses the core limitations of traditional ISH, enabling researchers to accurately visualize RNA localization and distribution within intact cells and tissues. This guide provides a detailed, evidence-based comparison of RNAscope performance against traditional ISH alternatives, supported by experimental data and validated protocols.
Technical Comparison: Mechanism and Performance
Probe Design and Specificity
The critical difference between RNAscope and traditional ISH lies in the fundamental probe architecture and its mechanism for ensuring specificity.
- Traditional ISH: Employs single, long, linear probes (often 200-500 bases) that are prone to non-specific binding. Amplification of background noise occurs alongside target signal, making it difficult to distinguish true expression from artifact [1] [2].
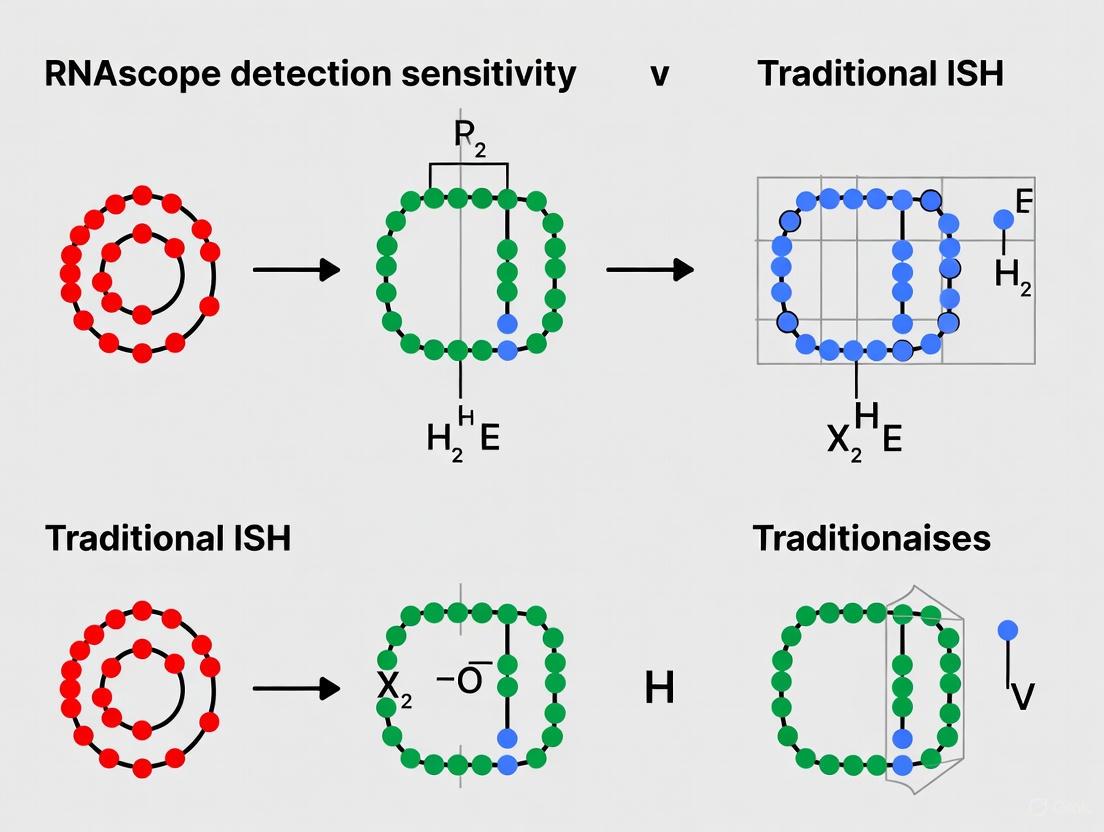
- RNAscope: Utilizes a proprietary "double Z" probe design. Each target is detected by ~20 pairs of probes. For signal amplification to occur, two "Z" probes must bind immediately adjacent to each other on the target RNA. This requirement makes it statistically improbable for non-specifically bound probes to generate a false-positive signal, thereby dramatically suppressing background [4] [1] [2].
The following diagram illustrates this fundamental mechanism that enables RNAscope's high specificity:
Quantitative Performance Metrics
Robust experimental data from peer-reviewed studies demonstrates the superior performance of RNAscope across key metrics. The table below summarizes a direct, quantitative comparison:
Table 1: Performance Comparison: RNAscope vs. Traditional ISH
| Performance Metric | Traditional ISH | RNAscope Technology | Experimental Context & Citation |
|---|---|---|---|
| Sensitivity | Limited detection of low-copy RNAs; unable to reliably detect single molecules. | Single-molecule sensitivity; each punctate dot represents an individual RNA transcript [4] [5]. | Validation using control genes with known expression levels (e.g., PPIB, UBC) in FFPE tissues [6]. |
| Specificity | High background noise; non-specific binding common. | High specificity due to dual Z-probe design; minimal background [4] [2]. | Staining with bacterial DapB negative control probe typically results in score of 0 (no staining) [6]. |
| Signal-to-Noise Ratio | Low; signal amplification also amplifies background. | Exceptionally high; background suppression is integral to the probe design [1] [2]. | Direct visualization of clear, distinct dots against a clean background in both fluorescent and chromogenic assays [5]. |
| Reproducibility | Variable due to complex, technique-sensitive protocols. | High inter-laboratory and inter-operator reproducibility [3]. | Standardized, validated protocols for manual and automated platforms (Ventana, Leica) [6]. |
| Compatibility with FFPE | Challenging due to RNA cross-linking and degradation. | Excellent; optimized for formalin-fixed, paraffin-embedded (FFPE) tissues, even with partially degraded RNA [4] [2]. | Successful detection in archival breast cancer TMAs with varying tissue age and storage conditions [5]. |
Application in Challenging Research Scenarios
The performance advantages of RNAscope translate into tangible benefits for complex research applications. A prime example is in the field of cardiac regeneration research, where the unequivocal identification of cardiomyocyte (CM) nuclei is crucial for accurately identifying cycling CMs.
- Challenge: Traditional methods using antibodies against sarcomeric proteins were error-prone, with a reported sensitivity of only 43% and specificity of 89% for identifying CM nuclei. Even with wheat germ agglutinin (WGA) staining to outline membranes, sensitivity only reached 65% [7].
- RNAscope Solution: Researchers designed intronic RNAscope probes targeting Tnnt2 (a sarcomeric protein) pre-mRNA. These probes label intronic RNAs retained in the nucleus, enabling specific nuclear localization of CMs [7].
- Result: The Tnnt2 intronic probe showed high colocalization with a genetic nuclear label (Obscurin-H2B-GFP), demonstrating CM specificity. It reliably labeled CM nuclei throughout all stages of mitosis, even after nuclear envelope breakdown, facilitating the investigation of CM dynamics after myocardial infarction [7]. This approach overcame the shortcomings of antibody-based methods and genetically modified mouse models.
Experimental Validation and Workflow
Detailed Protocol for Validation
To ensure reliable results, a standardized workflow for assay qualification is recommended. The key steps for validating the RNAscope assay for a new target or tissue type are outlined below, incorporating both the recommended protocol and experimental data from recent studies:
Table 2: Essential Research Reagent Solutions for RNAscope Assay Validation
| Reagent / Material | Function / Purpose | Critical Usage Notes |
|---|---|---|
| Positive Control Probes (e.g., PPIB, POLR2A, UBC) | Assess sample RNA integrity and assay performance. | PPIB staining should generate a score ≥2; required for qualifying sample [6]. |
| Negative Control Probe (dapB) | Assess background and non-specific signal. | Should yield a score of <1 (no staining or <1 dot/10 cells) [6]. |
| HybEZ Hybridization System | Maintains optimum humidity and temperature (40°C) during hybridization. | Required for manual assays; critical for consistent results [6]. |
| Superfrost Plus Slides | Provide superior tissue adhesion. | Other slide types may result in tissue detachment during the procedure [6]. |
| Immedge Hydrophobic Barrier Pen | Creates a well around the tissue section to retain reagents. | Specific brand required; others may fail during the assay [6]. |
| Specific Mounting Media (e.g., EcoMount, PERTEX, CytoSeal XYL) | Preserves staining for microscopy. | Media is assay-specific; using incorrect media can degrade signal [6]. |
The following diagram summarizes the key steps and decision points in the experimental workflow for a successful RNAscope assay:
Data Analysis and Quantification
A significant advantage of RNAscope is the objective, quantitative nature of its output. The standard scoring system is based on counting discrete, punctate dots per cell, which correlates directly with RNA copy number [6].
- Scoring Guidelines:
- Score 0: No staining or <1 dot/10 cells.
- Score 1: 1-3 dots/cell (visible at 20-40x magnification).
- Score 2: 4-9 dots/cell; very few dot clusters.
- Score 3: 10-15 dots/cell; <10% dots in clusters.
- Score 4: >15 dots/cell; >10% dots in clusters [6].
Advanced computational image analysis tools further enhance quantification. For instance, QuantISH is an open-source pipeline that automates cell segmentation and RNA dot quantification in chromogenic RNAscope (RNA-CISH) images, increasing reproducibility and efficiency over manual counting [8]. More recently, deep learning segmentation networks have been developed that can even outperform manual expert annotation in accurately identifying RNAscope dot positions, achieving a higher F1-score (0.745 for the model vs. 0.596 for inter-rater agreement) [5].
The evidence from experimental data and peer-reviewed studies consistently demonstrates that RNAscope technology effectively overcomes the fundamental challenge of RNA instability and the limitations of legacy ISH. Its proprietary double Z probe design provides a foundational advantage, enabling single-molecule sensitivity and exceptional specificity that traditional methods cannot reliably achieve.
This performance is not merely technical but translates into practical research benefits: enabling the use of challenging FFPE samples, providing objective quantification, and unlocking new research avenues—such as the identification of cardiomyocyte nuclei with intronic probes—that were previously hampered by technical constraints. For researchers and drug development professionals requiring precise, reliable, and spatially resolved RNA expression data, RNAscope represents a robust and validated solution that sets a new standard for in situ RNA analysis.
The field of RNA in situ hybridization (ISH) has undergone a revolutionary transformation, evolving from traditional methods reliant on radioactive probes to advanced signal amplification technologies that enable single-molecule detection with remarkable specificity. This evolution addresses a critical challenge in molecular pathology: the accurate visualization of RNA biomarkers within their native tissue context. Traditional RNA ISH techniques have been hampered by technical complexity, insufficient sensitivity, and limited specificity, restricting their clinical application to highly expressed targets like Epstein-Barr virus transcripts [9]. The introduction of novel ISH technologies represents a fundamental advancement in spatial genomics, bringing the benefits of in situ analysis to a wide range of RNA biomarkers and enabling rapid development of RNA ISH-based molecular diagnostic assays.
This comparison guide objectively examines the performance of the RNAscope platform against traditional ISH methods and emerging alternatives, providing researchers and drug development professionals with experimental data and protocols to inform their technology selection. By framing this analysis within the broader thesis of detection sensitivity evolution, we highlight how sophisticated probe design and amplification strategies have overcome historical limitations, creating new opportunities for biomarker discovery and validation.
Technical Comparison: Mechanism and Performance
Fundamental Technological Differences
The core distinction between traditional ISH and modern amplification technologies lies in their probe design and signal generation mechanisms. Traditional ISH typically utilizes single, directly labeled probes that hybridize to target RNA sequences. This approach generates limited signal and suffers from significant background noise due to non-specific hybridization, particularly challenging for low-abundance RNA targets [9].
In contrast, RNAscope employs a patented double-Z probe design where pairs of target probes must hybridize contiguously to a target region (approximately 50 bases) to create a binding site for subsequent amplification molecules [4] [9]. Each target probe contains a target-binding region, spacer sequence, and tail sequence. The two tail sequences together form a 28-base hybridization site for the pre-amplifier, which then binds multiple amplifiers and label probes [9]. This design ensures that single probes binding nonspecifically cannot trigger amplification, providing exceptional background control [4].
BaseScope, a variant for shorter targets, utilizes a similar principle but is optimized to detect RNA targets as short as 50-300 nucleotides, including splicing variants, point mutations, small insertions or deletions [10]. Both technologies represent a significant departure from traditional ISH through their requirement for probe pairing to initiate signal generation.
Performance Metrics and Experimental Data
Comparative studies and technical specifications demonstrate substantial performance differences between these technologies. The table below summarizes key performance metrics based on experimental data:
Table 1: Performance Comparison of RNA Detection Technologies
| Technology | Detection Sensitivity | Probe Design | Background Control | Target Length Requirements | Compatible Sample Types |
|---|---|---|---|---|---|
| Traditional ISH | Limited for low-copy targets | Single linear probes | Moderate to high due to non-specific hybridization | Typically >300 nucleotides | FFPE, frozen tissues, cells |
| RNAscope | Single-molecule detection [9] | Double-Z probes (~20 pairs per target) [4] | Exceptional; specific amplification requires paired probe binding [4] | 300-1000+ nucleotides [9] | FFPE, frozen tissues, cells, PBMCs [11] |
| BaseScope | High for short targets | Modified Z-probes | Excellent; maintains proprietary background suppression | 50-300 nucleotides [10] | FFPE tissues, cell cultures [10] |
Quantitative data from application studies further validates these performance characteristics. In foot-and-mouth disease virus detection, BaseScope demonstrated reliability despite tissue storage variations, maintaining detection sensitivity even when tissues were stored in formalin for up to 7 days and cut sections stored for up to 3 months [10]. RNAscope studies consistently show detection of low-copy reference genes (e.g., PPIB at 10-30 copies/cell) with scores ≥2, and high-copy genes (e.g., UBC) with scores ≥3, while negative controls (bacterial dapB) typically score <1, indicating minimal background [6].
Experimental Protocols and Workflows
RNAscope Assay Protocol
The RNAscope assay workflow comprises standardized steps that maintain consistency across applications. For formalin-fixed, paraffin-embedded (FFPE) tissues, the protocol begins with baking slides at 60°C for 1-2 hours, followed by deparaffinization with xylene and ethanol washes [9]. Critical pretreatment steps include:
- Target Retrieval: Boiling in citrate buffer (10 mmol/L, pH 6) for 15 minutes [9]
- Protease Digestion: Treatment with protease (10 μg/mL) at 40°C for 30 minutes in a HybEZ hybridization oven [9]
Following pretreatment, the core hybridization and amplification steps proceed:
- Probe Hybridization: Incubate with target probes in hybridization buffer at 40°C for 2 hours [6]
- Amplification Cascade:
- Preamplifier hybridization (30 minutes at 40°C)
- Amplifier hybridization (15 minutes at 40°C)
- Label probe hybridization (15 minutes at 40°C) [9]
- Signal Detection: Chromogenic development with DAB or Fast Red, followed by counterstaining with hematoxylin [9]
The entire manual procedure can be completed in 7-8 hours or conveniently divided over two days [6]. The protocol is also adaptable for automated staining systems including Ventana DISCOVERY XT/ULTRA and Leica BOND RX [12].
BaseScope Protocol Modifications
The BaseScope assay follows a similar workflow but with specific optimizations for shorter targets. Critical adjustments identified in foot-and-mouth disease virus detection include strict adherence to prescribed reagents and equipment, as attempts to use alternatives were unsuccessful, demonstrating the "fastidious nature of this diagnostic modality" [10]. The synergistic characteristics of commercial assay components proved essential for successful detection.
Traditional ISH Protocol Limitations
Traditional ISH protocols typically involve longer hybridization times (often overnight), less standardized pretreatment conditions, and lack the structured amplification steps that characterize modern methods. Without built-in background suppression mechanisms, they require extensive optimization and often yield inconsistent results across laboratories [9].
Advanced Applications and Integration Approaches
Multimodal Integration with Complementary Techniques
Advanced research applications increasingly combine RNA ISH technologies with other detection modalities to gain comprehensive biological insights:
RNAscope-IHC Co-detection: A protocol for combining RNAscope and immunohistochemistry enables simultaneous visualization of inflammatory gene products in neurons and microglia [13]. This approach uses RNAscope to label transcripts of interest and IHC to label cell-type specific antigens (e.g., IBA1 for microglia, NeuN for neurons), allowing quantification of RNA transcripts within specific cell boundaries using confocal microscopy [13].
RNA-seq Correlation: Studies analyzing nine cancer biomarkers (ESR1, PGR, ERBB2, CD274, etc.) across 365 FFPE samples revealed strong correlations between RNA-seq and IHC data, with coefficients ranging from 0.53 to 0.89 [14]. This demonstrates how RNA expression thresholds can be established to reflect protein expression classifications, enabling cross-platform validation.
Computational Analysis Frameworks
The development of automated image analysis tools has enhanced the quantitative potential of modern ISH technologies:
QuantISH: An open-source RNA-ISH image analysis framework quantifies marker expressions in individual carcinoma, immune, and stromal cells from chromogenic or fluorescent ISH images [8]. This pipeline performs cell segmentation, cell-type classification based on nuclear morphology, and expression quantification at single-cell resolution, enabling analysis of tumor heterogeneity [8].
HALO Software: Commercial platforms provide automated quantification of RNAscope signals, counting dot numbers per cell to determine RNA copy numbers [12].
Essential Research Reagent Solutions
Successful implementation of advanced ISH technologies requires specific reagents and equipment. The following table details essential components for establishing these capabilities in research laboratories:
Table 2: Essential Research Reagents and Equipment for Advanced ISH Applications
| Item Category | Specific Product/Requirement | Function/Purpose | Technology Compatibility |
|---|---|---|---|
| Slide System | Superfrost Plus slides | Prevents tissue detachment during stringent processing | RNAscope, BaseScope [6] |
| Hybridization System | HybEZ Oven | Maintains optimum humidity and temperature during hybridization | RNAscope [6] [15] |
| Control Probes | PPIB, POLR2A (positive); dapB (negative) | Assess sample RNA quality and optimal permeabilization | RNAscope, BaseScope [6] [11] |
| Detection Kits | RNAscope 2.5 HD Brown/Red Reagent Kit | Provides amplification and detection reagents | RNAscope [6] |
| Barrier Pen | ImmEdge Hydrophobic Barrier Pen | Maintains hydrophobic barrier throughout procedure | RNAscope [6] |
| Mounting Media | EcoMount, PERTEX (for fluorescent assays) | Preserves signal without quenching fluorescence | RNAscope [6] |
| Image Analysis | HALO Software, QuantISH | Quantifies signals per cell automatically | RNAscope, BaseScope [12] [8] |
The evolution from radioactive probes to sophisticated signal amplification technologies represents a quantum leap in RNA detection capabilities. RNAscope and BaseScope technologies have demonstrated superior performance in sensitivity, specificity, and reliability compared to traditional ISH methods, enabling researchers to address previously intractable questions in gene expression analysis.
The double-Z probe design philosophy underlying these advanced platforms provides a fundamental advantage: the requirement for paired probe binding before signal amplification virtually eliminates background noise while maintaining single-molecule sensitivity. This technical breakthrough, combined with standardized workflows and specialized reagents, has transformed spatial transcriptomics from a specialized technique to an accessible, robust tool for both basic research and clinical applications.
As these technologies continue to evolve and integrate with complementary approaches like IHC and RNA-seq, they promise to further enhance our understanding of gene expression patterns in their native tissue context, accelerating biomarker discovery and therapeutic development across diverse disease areas.
The advent of RNAscope technology represents a paradigm shift in the field of RNA in situ hybridization (ISH), addressing long-standing challenges of sensitivity and specificity that plagued traditional methods. This guide details the core breakthrough—the novel 'Double Z' probe design—that enables single-molecule RNA visualization within morphologically intact tissues. We objectively compare its performance against traditional ISH and other modern techniques, supported by experimental data and detailed methodologies, providing researchers and drug development professionals with a clear understanding of its capabilities and applications in spatial genomics.
The Fundamental Challenge in Traditional RNA ISH
Traditional RNA in situ hybridization techniques have been limited by a fundamental trade-off between sensitivity and specificity. Methods using single-labeled probes, such as those labeled with digoxigenin or fluorophores, often struggled with high background noise resulting from non-specific probe binding [16]. This background problem was exacerbated when signal amplification was attempted, as amplification systems could not distinguish between true target binding and non-specific background, thus amplifying both equally [1]. Consequently, the sensitivity of traditional RNA ISH remained insufficient for detecting low-abundance RNA transcripts, restricting its utility to highly expressed targets such as Epstein-Barr virus-encoded RNA (EBER) in clinical settings [9]. The spatial context of gene expression, crucial for understanding cellular heterogeneity and function in complex tissues, was often lost in "grind-and-bind" methods like RT-PCR that required RNA extraction [9].
The RNAscope Solution: 'Double Z' Probe Design Explained
Core Architecture and Mechanism
RNAscope's revolutionary approach centers on a patented double-Z probe design that fundamentally reengineers the hybridization process [9] [4]. This system employs pairs of target probes ("Z probes") that must bind in tandem to the RNA target for signal amplification to initiate [9]. Each individual Z probe contains three distinct elements:
- Lower Region: An 18-25 base sequence complementary to the target RNA [4]
- Spacer Sequence: A linker connecting the target-complementary region to the tail [9]
- Tail Sequence: A 14-base segment that, when paired with an adjacent probe, creates a 28-base binding site for the pre-amplifier [9]
The critical innovation is that both probes must bind contiguously (~50 base region) to the target RNA to form the complete recognition site for the pre-amplifier molecule [9] [1]. This dual-binding requirement provides exceptional specificity, as it is statistically improbable that two independent probes would bind nonspecifically to off-target sequences in immediate juxtaposition [4].
Signal Amplification Cascade
Once the double-Z probe pair successfully hybridizes to the target RNA, a multi-step amplification cascade ensues:
- Preamplifier Binding: The 28-base site formed by the Z-probe tails recruits the preamplifier molecule [9]
- Amplifier Hybridization: Each preamplifier contains 20 binding sites for amplifier molecules [9]
- Label Probe Attachment: Each amplifier provides 20 sites for label probes conjugated with fluorescent dyes or chromogenic enzymes [9]
This branching amplification system theoretically generates up to 8,000 labels for each target RNA molecule when 20 probe pairs are used [9], providing exceptional detection sensitivity while maintaining specificity through the initial dual-probe requirement.
Visualization of RNAscope's Signal Amplification Cascade. The double-Z probe design creates a specific binding site that initiates a branching amplification pathway, enabling detection of individual RNA molecules.
Specialized Probe Variations for Diverse Applications
The core double-Z technology has been adapted into specialized assay systems optimized for different molecular targets:
Table 1: RNAscope Technology Portfolio for Different Target Types
| Assay Type | Target Length | Probe Pairs | Primary Applications |
|---|---|---|---|
| RNAscope | >300 nucleotides [17] | ~20 ZZ pairs [18] | mRNA, long non-coding RNA, viral RNA [19] |
| BaseScope | 50-300 nucleotides [17] | 1-3 ZZ pairs [18] | Short targets, splice variants, point mutations [1] |
| miRNAscope | 17-50 nucleotides [17] | Proprietary design | microRNAs, siRNAs, ASOs [18] |
Experimental Validation: Protocols and Performance Data
Key Methodological Framework
The RNAscope assay workflow follows a structured protocol that maintains RNA integrity while enabling specific probe access:
Sample Preparation: Tissues are fixed in 10% neutral buffered formalin (NBF) for 16-32 hours at room temperature, then processed for paraffin embedding or frozen sectioning [17]. Section thickness recommendations: 5±1 μm for FFPE, 10-20 μm for fresh frozen tissues [17].
Pretreatment Protocol: Slides undergo heat-induced epitope retrieval in citrate buffer (100-103°C for 15 minutes) followed by protease digestion (10 μg/mL at 40°C for 30 minutes) to permeabilize tissues and unmask target RNA [9].
Hybridization and Amplification: Target probes are hybridized at 40°C for 3 hours in a specialized HybEZ oven, followed by sequential 30-minute incubations with preamplifier, amplifier, and label probes [9].
Controls and Validation: Each experiment includes positive control probes (housekeeping genes PPIB, POLR2A, or UBC) and negative control probes (bacterial dapB gene) to verify RNA quality and assay specificity [17] [6].
Quantitative Performance Comparison
Table 2: Experimental Comparison Between RNA ISH Platforms
| Performance Metric | Traditional RNA ISH | RNAscope | Validation Method |
|---|---|---|---|
| Sensitivity | Limited to highly expressed genes [9] | Single-molecule detection [9] | QuantiGene 2.0 correlation [20] |
| Specificity | High background, non-specific binding [16] | 100% specificity claimed [16] | Negative control (dapB) verification [17] |
| Concordance with qPCR | Not reliably established | 81.8-100% [16] | Systematic review comparison [16] |
| Concordance with IHC | Variable, target-dependent | 58.7-95.3% [16] | Parallel staining assessment [16] |
| Single-Cell Resolution | Limited by background noise | Precise transcript counting [4] | Manual dot counting vs. digital quantification [6] |
Experimental Evidence for Single-Molecule Sensitivity
The original validation studies demonstrated RNAscope's single-molecule sensitivity through multiple approaches:
- HER2 DNA Detection: In HeLa cells, RNAscope consistently showed two fluorescent puncta per cell, corresponding precisely to the diploid status of the HER2 gene [20]
- Transcript Quantification: Manual counting of HER2 mRNA puncta in HeLa cells showed excellent correlation with transcript numbers measured by QuantiGene 2.0 assay [20]
- Signal Intensity Analysis: The fluorescence intensity of individual HER2 mRNA puncta matched that of HER2 DNA signals, confirming single-transcript detection [20]
Comparative Analysis with Alternative Technologies
RNAscope vs. Traditional ISH and IHC
When compared with traditional RNA ISH methods, RNAscope demonstrates superior performance across multiple parameters. Traditional ISH techniques using digoxigenin-labeled or radioactive probes produce diffuse signals with considerable background, limiting accurate quantification [1]. In contrast, RNAscope generates discrete punctate signals where each dot represents an individual RNA molecule, enabling precise transcript counting and localization [21].
Compared to immunohistochemistry (IHC), RNAscope offers advantages for targets where antibodies are unavailable or perform poorly. The systematic review by PMC8710359 found that while RNAscope showed good concordance with IHC for many targets (58.7-95.3%), it could detect low-expression targets not visible by IHC and provided more objective, quantifiable signals [16] [21]. This makes it particularly valuable for detecting secreted proteins, cytokines, and low-abundance targets where antibody-based detection struggles [19].
RNAscope vs. PCR-Based Methods
While quantitative RT-PCR remains the gold standard for bulk RNA quantification, it destroys tissue architecture and cannot provide spatial context [9]. RNAscope bridges this gap by enabling in situ quantification while preserving morphological information. The technology shows high concordance with qPCR (81.8-100%) while offering the critical advantage of cellular localization [16]. This spatial dimension is particularly valuable for heterogeneous tissues like tumors, where the cellular origin of gene expression significantly impacts biological interpretation.
Implementation Guide: Research Reagent Solutions
Table 3: Essential Research Reagents and Equipment for RNAscope
| Component | Specification | Function | Critical Notes |
|---|---|---|---|
| Probe Sets | Target-specific ZZ designs [18] | Target RNA hybridization | 20 ZZ pairs for standard RNAscope; 1-3 for BaseScope [18] |
| Control Probes | PPIB, POLR2A, UBC (positive); dapB (negative) [17] | Assay validation | Essential for interpreting results; species-specific [6] |
| HybEZ Oven | Temperature and humidity control system [17] | Hybridization incubation | Maintains 40°C with proper humidity; required for manual assays [17] |
| Detection Reagents | Chromogenic (DAB, Fast Red) or fluorescent [9] | Signal visualization | Fluorophores for multiplexing; enzymes for bright-field microscopy [9] |
| Slide Type | SuperFrost Plus [17] | Tissue adhesion | Prevents tissue detachment during stringent washes [17] |
| Barrier Pen | ImmEdge Hydrophobic Barrier Pen [17] | Reagent containment | Maintains barrier throughout procedure; other pens may fail [17] |
Advanced Applications and Multiplexing Capabilities
The RNAscope platform supports sophisticated experimental designs including multiplex detection of up to four RNA targets simultaneously using spectrally distinguishable fluorescent labels [9]. For higher-plex applications, the HiPlex system enables detection of up to 12 targets through sequential hybridization and signal removal [19]. Furthermore, RNAscope is compatible with simultaneous protein detection via immunohistochemistry on the same tissue section, enabling integrated analysis of RNA and protein expression within identical cellular contexts [20]. This multiomic capability is particularly powerful for investigating gene regulatory networks, validating therapeutic targets, and characterizing complex tissue microenvironments in fields such as immuno-oncology and neuroscience.
The technology has proven particularly valuable in gene and cell therapy development, enabling researchers to visualize AAV vector biodistribution, transgene expression, CAR-T cell infiltration, and cellular tropism with spatial context [19]. These applications demonstrate how the core double-Z probe design has evolved into a versatile platform addressing diverse research needs across biological disciplines.
RNAscope's double-Z probe design represents a fundamental advancement in RNA detection technology, successfully addressing the critical limitations of traditional ISH methods. Through its requirement for contiguous probe binding and branched signal amplification, it achieves unprecedented specificity and sensitivity enabling single-molecule visualization. While traditional techniques remain valuable for specific applications, RNAscope provides researchers with a powerful spatial genomics tool that bridges the gap between bulk RNA measurement and cellular context. As the field moves toward increasingly multiplexed and multiomic analyses, the principles embodied in the double-Z design continue to enable new possibilities for understanding gene expression in situ.
In situ hybridization (ISH) has long been a fundamental tool for visualizing gene expression within the morphological context of intact tissues. However, traditional ISH methods have been plagued by significant limitations, including poor sensitivity, high background noise, and an inability to reliably detect low-abundance transcripts. These technical constraints have hindered researchers' ability to study gene expression with the precision required for modern molecular biology and drug development applications. The advent of RNAscope Technology represents a paradigm shift in RNA ISH, introducing a novel signal amplification and background suppression system that enables single-molecule visualization of RNA transcripts as distinct, quantifiable dots within individual cells.
This revolutionary approach transforms RNA detection from a qualitative assessment to a precise, quantitative science. Where traditional methods might show diffuse staining or ambiguous signals, RNAscope generates discrete, easily distinguishable dots, with each dot corresponding to a single RNA molecule. This single-molecule advantage provides researchers and drug development professionals with unprecedented accuracy in gene expression analysis, enabling more confident interpretation of results and more reliable decision-making in therapeutic development pipelines.
Technology Comparison: RNAscope Versus Traditional ISH
Fundamental Mechanism Differences
The superior performance of RNAscope compared to traditional ISH stems from fundamental differences in their detection methodologies. Traditional ISH relies on single linear probes that directly hybridize to target RNA, with detection typically involving simple antibody-based systems that yield diffuse signals and considerable background noise. This approach struggles with sensitivity and specificity, particularly for low-abundance transcripts.
In contrast, RNAscope employs a patented double-Z probe design that creates a unique amplification architecture. Each RNAscope probe pair consists of two separate oligonucleotides that bind side-by-side to the target RNA, forming a docking site for pre-amplifier molecules. This design enables substantial signal amplification while simultaneously incorporating background suppression through a proprietary method that minimizes non-specific binding. The result is the ability to visualize individual RNA molecules as distinct dots with single-molecule resolution, a capability unattainable with conventional ISH methods [22].
Table 1: Comparative Analysis of Traditional ISH vs. RNAscope Technology
| Feature | Traditional ISH | RNAscope Technology |
|---|---|---|
| Probe Design | Single linear probes | Patented double-Z probe pairs |
| Signal Amplification | Limited or none | Multistep amplification system |
| Background Suppression | Minimal, high non-specific binding | Built-in background suppression |
| Detection Sensitivity | ~20-30 copies/cell | Single-molecule detection |
| Quantification Capability | Qualitative or semi-quantitative | Semi-quantitative (dots=copies) |
| Resolution | Cellular | Single-molecule |
| Signal Appearance | Diffuse staining | Discrete, distinct dots |
| Validation Requirements | Extensive optimization | Standardized controls |
Performance Metrics and Experimental Validation
Experimental data from numerous studies demonstrates the remarkable advantage of RNAscope over traditional ISH. The technology achieves single-molecule sensitivity while maintaining exceptional specificity, with each visualized dot corresponding to an individual RNA transcript. This quantification capability enables researchers to precisely monitor gene expression changes in response to experimental conditions or therapeutic interventions.
The scoring system developed for RNAscope highlights its quantitative nature, with established guidelines that correlate dot counts per cell with transcript abundance: a score of 0 indicates no staining or <1 dot/10 cells; score 1 represents 1-3 dots/cell; score 2 indicates 4-9 dots/cell; score 3 represents 10-15 dots/cell; and score 4 indicates >15 dots/cell with >10% dots in clusters [6]. This systematic approach to quantification stands in stark contrast to the subjective interpretation often required with traditional ISH methods.
Recent applications in challenging research areas further demonstrate RNAscope's capabilities. In cardiac regeneration studies, intronic RNAscope probes have enabled precise identification of cardiomyocyte nuclei based on nascent transcript detection, overcoming limitations of antibody-based methods that struggle with specificity and nuclear localization during cell division [23]. Similarly, the BaseScope assay, a variant optimized for shorter targets, has proven effective for detecting foot-and-mouth disease virus in carrier animals, demonstrating that even short RNA targets (50-300 nucleotides) can be reliably detected with single-molecule precision [10].
Experimental Protocols and Methodologies
Standardized RNAscope Workflow
The RNAscope assay procedure follows a meticulously optimized protocol that can be completed in 7-8 hours, either in a single day or conveniently divided over two days. Unlike traditional ISH that requires RNase-free conditions, RNAscope assays do not need an RNase-free environment, significantly simplifying laboratory workflow [6]. Most reagents are available in convenient Ready-To-Use (RTU) dropper bottles, creating a nearly pipette-free workflow that enhances reproducibility and ease of use.
Critical steps in the protocol include:
- Sample Preparation: Fixation in fresh 10% neutral buffered formalin (NBF) for 16-32 hours followed by standard processing and embedding
- Antigen Retrieval: Heat-induced epitope retrieval under standardized conditions
- Protease Digestion: Controlled protease treatment to permeabilize tissue while preserving RNA integrity
- Hybridization: Probe hybridization at 40°C using the HybEZ System to maintain optimum humidity and temperature
- Signal Amplification: Sequential amplifier applications that build the detection complex
- Detection: Chromogenic or fluorescent detection of the amplified signal
For challenging samples where preparation conditions are unknown or suboptimal, ACD recommends a qualification workflow using control probes including housekeeping genes (PPIB, POLR2A, or UBC) and negative control (bacterial dapB) to verify sample RNA quality and optimal permeabilization before attempting target gene detection [6].
Specialized Applications and Modifications
The versatility of the RNAscope platform enables adaptation for specialized research needs. The BaseScope assay provides enhanced sensitivity for shorter targets, including RNA splicing variants, point mutations, small insertions or deletions, and short RNA targets of 50-300 nucleotides [10]. This capability is particularly valuable for detecting viral RNAs, SNP variants, and alternatively spliced transcripts that would be undetectable with traditional ISH.
For multiplex applications, RNAscope technology supports simultaneous detection of multiple RNA targets through either chromometric or fluorescent detection systems. The newly enhanced RNAscope Multiplex Fluorescent V2 Assay with Fluorescent TSA Vivid Dyes expands the color palette available for researchers needing to visualize multiple targets in the same sample [24]. This advanced multiplexing capability enables complex co-localization studies and pathway analysis within the morphological context of intact tissue.
Recent innovations include HiPlex assays that expand visualization to 8 or 12 probes in the same sample, dramatically increasing the amount of information that can be gathered from a single tissue section [22]. Additionally, combination assays that enable simultaneous detection of RNA and protein biomarkers (RNA-protein co-detection) provide a comprehensive view of gene expression and regulation at the cellular level.
Research Reagent Solutions and Essential Materials
Successful implementation of RNAscope technology requires specific reagents and equipment optimized for the assay system. The fastidious nature of this diagnostic modality was demonstrated in a study where attempts to use in-house reagents or equipment as alternatives to the prescribed workflow were unsuccessful, highlighting the synergistic characteristics of this commercial assay [10].
Table 2: Essential Research Reagents and Materials for RNAscope Experiments
| Reagent/Material | Specification | Function | Importance |
|---|---|---|---|
| Probes | Catalog (13,000+ targets) or custom designs | Target-specific hybridization | Species-specific binding with proprietary Z-probe design |
| Superfrost Plus Slides | Fisher Scientific Cat. No. 12-550-15 | Tissue adhesion | Prevents tissue detachment during stringent washes |
| ImmEdge Hydrophobic Barrier Pen | Vector Laboratories Cat. No. 310018 | Creates hydrophobic barrier around tissue | Prevents reagent evaporation and tissue drying |
| HybEZ Hybridization System | ACD Proprietary System | Humidity and temperature control | Maintains optimal hybridization conditions |
| Detection Reagents | Chromogenic or fluorescent | Signal development | Enzyme-based detection with high sensitivity |
| Mounting Media | Xylene-based (Brown) or EcoMount/PERTEX (Red) | Slide preservation | Preserves signal and tissue morphology |
The proprietary double-Z probe design forms the foundation of RNAscope's performance advantages. Each probe pair consists of two separate oligonucleotides that bind side-by-side on the target RNA, creating a docking site for pre-amplifier molecules. This design enables dramatic signal amplification while preventing non-specific background because the full amplifier complex can only assemble when both probe fragments are correctly hybridized to their target in close proximity [22]. The probe catalog includes over 13,000 probes from more than 140 species and over 100 viruses, with custom probes also available upon request to meet specialized research needs.
Visualization and Data Interpretation
Signal Pattern Recognition and Quantification
The discrete dot-like pattern of RNAscope signals represents a significant advantage for data interpretation compared to the diffuse staining typical of traditional ISH. Each dot corresponds to an individual RNA molecule, enabling direct quantification of transcript abundance at the cellular level. This clear signal pattern eliminates much of the subjectivity associated with traditional ISH interpretation and provides researchers with quantifiable data for statistical analysis.
Proper scoring follows established guidelines that focus on counting dots per cell rather than assessing signal intensity. The semi-quantitative scoring system ranges from 0 (no staining or <1 dot/10 cells) to 4 (>15 dots/cell with >10% dots in clusters), providing a standardized framework for comparing expression levels across experimental conditions [6]. For genes with expression levels outside the typical range of the control probes (PPIB), the scoring criteria may need to be scaled accordingly to accurately capture expression differences.
Troubleshooting and Optimization Strategies
While RNAscope technology offers robust performance, proper optimization is essential for challenging samples. Common issues include:
- No Signal: Confirm positive control probes work as expected; verify sample RNA quality; check protease concentration and incubation time
- High Background: Ensure proper protease digestion; verify reagent freshness; confirm appropriate probe concentrations
- Tissue Detachment: Use only Superfrost Plus slides; verify fixation conditions; avoid drying between steps
- Weak Signal: Optimize antigen retrieval; extend protease digestion for over-fixed tissues; check probe hybridization efficiency
For automated platforms (Ventana DISCOVERY XT/ULTRA or Leica BOND RX), specific instrument maintenance is critical, including regular decontamination to prevent microbial growth in fluidic lines and replacement of bulk solutions with recommended buffers before running RNAscope assays [6]. The Leica BOND RX system offers flexible pretreatment options, with standard conditions of 15 minutes Epitope Retrieval 2 (ER2) at 95°C and 15 minutes Protease at 40°C, which can be adjusted in increments for over-fixed tissues or challenging samples.
Application Case Studies in Research and Drug Development
Advanced Research Applications
The single-molecule detection capability of RNAscope has enabled breakthroughs across multiple research domains. In cardiac regeneration research, intronic RNAscope probes have solved the critical challenge of unequivocally identifying cardiomyocyte nuclei in cardiac sections, a prerequisite for accurately identifying cycling cardiomyocytes. The Tnnt2 intronic RNAscope probe demonstrated high specificity for cardiomyocyte nuclei and remained closely associated with cardiomyocyte chromatin through all stages of mitosis, even during nuclear envelope breakdown—a period when nuclear protein markers become undetectable [23]. This capability has proven invaluable for investigating dynamics of DNA synthesis and potential mitoses in cardiomyocytes after myocardial infarction.
In drug development, RNAscope technology enables precise visualization and quantification of oligonucleotide therapy delivery, spatial biodistribution, and efficacy. The platform can detect various oligonucleotide drugs including ASOs, siRNAs, miRNAs, and aptamers, allowing researchers to assess on-target and off-target effects within intact tissues [25]. This application provides crucial insights during preclinical development by revealing the spatial distribution and functional efficacy of RNA drug candidates alongside endogenous RNA and protein markers.
Future Directions and Technology Evolution
The continued evolution of RNAscope technology points toward increasingly multiplexed assays and integration with complementary techniques. The recently introduced HiPlex assay system enables simultaneous detection of up to 12 different RNA targets in the same tissue section, dramatically increasing the information yield from precious clinical samples [22]. This advancement supports comprehensive pathway analysis and cell typing within the tissue microenvironment.
Integration with protein detection methods creates a powerful multiomics platform that enables researchers to correlate transcriptional activity with protein expression and post-translational modifications within the same cellular context. Additionally, automated image analysis solutions, such as Indica Labs' HALO platform with specialized RNAscope modules, facilitate high-throughput quantification of RNA expression patterns in large sample cohorts, supporting robust statistical analysis and biomarker discovery [26].
Diagram 1: RNAscope single-molecule detection advantage. The proprietary double-Z probe design enables specific signal amplification with background suppression, transforming diffuse signals into discrete, quantifiable dots that provide single-molecule resolution and enable accurate quantification and precise localization for research applications.
RNAscope technology represents a fundamental advancement in RNA detection methodology, offering researchers and drug development professionals unprecedented capability to visualize gene expression with single-molecule precision. The transformation of RNA signals from diffuse staining to discrete, quantifiable dots addresses long-standing limitations of traditional ISH and enables new applications in basic research, biomarker discovery, and therapeutic development. As the technology continues to evolve with enhanced multiplexing capabilities, improved detection sensitivity, and expanded integration with complementary methods, its role in advancing spatial biology and precision medicine will undoubtedly grow. For researchers requiring definitive RNA visualization within morphological context, RNAscope's single-molecule advantage provides the clarity and confidence needed to move scientific understanding forward.
The analysis of RNA biomarkers within their histopathological context is highly desirable in molecular pathology and research. However, this endeavor faces significant challenges when working with formalin-fixed, paraffin-embedded (FFPE) tissues—the most widely archived material in pathology departments globally. Formalin fixation induces protein-RNA cross-linkages and causes RNA fragmentation through molecular modifications, while prolonged archival storage can lead to further RNA degradation [27] [28]. These pre-analytical variables have traditionally limited the utility of archival specimens for RNA analysis, particularly for retrospective studies.
RNAscope in situ hybridization (ISH) represents a technological advancement that addresses these limitations. Unlike traditional RNA ISH methods, which struggle with sensitivity and specificity issues, RNAscope employs a unique "double Z" probe design that enables simultaneous signal amplification and background suppression [16] [2]. This technical overview examines how RNAscope performs on FFPE and degraded samples compared to traditional alternatives, providing researchers with evidence-based guidance for implementing this technology in their experimental workflows.
The Fundamental Principle: RNAscope's "Double Z" Probe Design
RNAscope's superior performance stems from its patented probe design and signal amplification system. The technology uses pairs of "Z" probes that specifically bind to the target RNA sequence. Each probe consists of three elements: a target-hybridizing region, a linker sequence, and a tail that binds pre-amplifier molecules [16]. This design requires two probes to bind adjacent to each other on the target RNA before signal amplification can proceed, providing exceptional specificity by preventing non-specific background noise [2].
The amplification cascade begins once the "Z" probe dimers form on the target RNA. Pre-amplifier molecules attach to the probe tails, followed by multiple amplifiers that bind to each pre-amplifier. Finally, labeled probes conjugate to the amplifiers, resulting in up to 8,000-fold signal amplification [16]. Each detected RNA molecule appears as a distinct dot, enabling single-molecule visualization and quantification while preserving tissue morphology [2] [5].
Table: Comparison Between Traditional ISH and RNAscope Technology
| Feature | Traditional ISH | RNAscope Technology |
|---|---|---|
| Probe Design | Single probes conjugated with labels | Pairs of "Z" probes requiring dimerization |
| Signal Amplification | Limited or none | Multi-stage (up to 8,000x amplification) |
| Specificity Control | Minimal background suppression | Built-in background suppression through probe design |
| Sensitivity | Limited to highly expressed genes | Single-molecule detection capability |
| RNA Degradation Tolerance | Poor | Can detect partially fragmented RNA |
| Quantification Potential | Qualitative or semi-quantitative | Semi-quantitative with dot counting |
RNAscope Signal Amplification Pathway: This diagram illustrates the sequential binding process that enables RNAscope to achieve high sensitivity and specificity, particularly valuable for degraded RNA samples.
Critical Differences from Immunohistochemistry (IHC)
While RNAscope shares some workflow similarities with IHC, several key differences impact its performance on FFPE samples. Unlike IHC, which detects proteins, RNAscope targets RNA molecules, requiring specific protease digestion steps to permeabilize tissue and expose target RNA [6]. Additionally, RNAscope requires specialized equipment like the HybEZ Hybridization System to maintain optimum humidity and temperature during hybridization steps [6]. Proper mounting media selection is also crucial, with specific requirements differing between the chromogenic red and brown assays [6].
Experimental Evidence: Performance on FFPE and Degraded Samples
Impact of Formalin Fixation Time
A systematic evaluation examined the effect of formalin-fixation time on RNAscope signal detection. Tissues were fixed in 10% neutral-buffered formalin for durations ranging from 1 to 270 days, with signal intensity and percent area measured for a reference gene (16S rRNA) [27]. The results demonstrated that signal intensity and percent area decreased significantly after 180 days of formalin fixation. While tissues maintained detectable signal at 180 days, no signal was observed at 270 days of continuous formalin fixation [27].
This degradation pattern aligns with the molecular changes in formalin-fixed tissues. During the initial 24-48 hours of fixation, formaldehyde forms reversible methylene glycol complexes that penetrate cells, creating protein cross-linkages. After approximately 30 days, covalent bonds form irreversibly, causing increased RNA damage through strand fragmentation and molecular modifications by adducts and cross-links [27].
Table: Effect of Prolonged Formalin Fixation on RNAscope Signal Intensity
| Fixation Time | Signal Intensity | Percent Area | Detection Capability |
|---|---|---|---|
| 1-28 days | Strong | High | Optimal detection |
| 60-90 days | Moderate | Moderate | Readily detectable |
| 180 days | Reduced but present | Reduced | Detectable with lower signal |
| 270 days | No signal | No signal | Not detectable |
Performance on Long-term Archival FFPE Blocks
To assess RNAscope's capability with archival materials, researchers compared detection of canine distemper virus (CDV) RNA via RNAscope with CDV antigen detection via immunohistochemistry in replicate sections from blocks stored at room temperature for intervals ranging from 6 months to 15 years [27]. Remarkably, RNA was detected in FFPE tissues stored for up to 15 years, demonstrating RNAscope's robustness for retrospective studies utilizing archival blocks [27].
A separate study on breast cancer samples further quantified RNA degradation over archival time, comparing FFPE tissues (FFPET) with fresh frozen tissues (FFT) [28]. As expected, RNAscope signals in FFPET were lower than in FFT in an archival duration-dependent fashion. The degradation was most pronounced in highly expressed housekeeping genes (UBC and PPIB) compared to low-to-moderate expressors (POLR2A and HPRT1) [28]. This finding highlights the importance of selecting appropriate control genes when working with archival samples.
Comparative Performance vs. Traditional Techniques
Systematic Review Evidence
A comprehensive systematic review evaluated RNAscope's performance in the clinical diagnostic field compared to established "gold standard" methods [16]. The review analyzed 27 retrospective studies, primarily focusing on cancer samples, with results showing that RNAscope has high concordance rates with qPCR, qRT-PCR, and DNA ISH (81.8-100%) [16]. However, the concordance with IHC was somewhat lower (58.7-95.3%), reflecting the different biomolecules measured by each technique (RNA vs. protein) [16].
This evidence suggests that RNAscope serves as a complementary technique alongside established methods rather than a wholesale replacement. It is particularly valuable for resolving ambiguous IHC results or when investigating genes for which reliable antibodies are unavailable [16] [28].
Advantages for Degraded Samples
RNAscope's unique probe design provides distinct advantages for analyzing degraded RNA in FFPE samples. Because the technique uses multiple short probe pairs targeting different regions of the same RNA molecule, it can detect partially fragmented RNA that might be missed by techniques requiring intact full-length molecules [16] [2]. This feature makes RNAscope particularly valuable for analyzing poorly preserved clinical specimens or archival tissues with unknown preservation histories.
The ability to visualize RNA within morphological context provides another advantage over bulk extraction methods like RT-qPCR. Even when RNA is partially degraded, researchers can still determine which cell types express the target RNA and how expression patterns relate to tissue pathology [28].
Essential Methodological Considerations for Suboptimal Samples
Quality Control Practices
Robust quality control is essential when working with FFPE and potentially degraded samples. ACD recommends a two-level quality control practice [29]:
Technical assay control: Verify proper assay performance using control samples with known housekeeping gene probes and negative control probes.
Sample/RNA quality control: Optimize pretreatment conditions for specific tissue samples using positive and negative control probes to confirm RNA integrity.
For quality assessment, researchers should utilize both negative control probes (targeting bacterial DapB gene to confirm absence of background staining) and appropriate positive control probes selected based on target expression level [29]:
- UBC (Ubiquitin C): For high expression targets (>20 copies/cell)
- PPIB (Cyclophilin B): Medium expression (10-30 copies/cell) - recommended for most tissues
- Polr2A (RNA polymerase II): Low expression targets (3-15 copies/cell)
Pretreatment Optimization for Suboptimal Samples
When sample preparation conditions are unknown or suboptimal, pretreatment optimization becomes crucial. The recommended workflow involves [6]:
- Running control slides with known positive and negative control probes
- Evaluating staining results using RNAscope scoring guidelines
- Adjusting pretreatment conditions based on control results before proceeding with experimental probes
For over-fixed tissues or samples with extended archival times, incremental adjustments to antigen retrieval and protease digestion times may be necessary. On automated platforms like the Leica BOND RX system, recommended adjustments include increasing ER2 time in 5-minute increments and protease time in 10-minute increments while maintaining constant temperatures [6].
Scoring and Quantification Approaches
Proper scoring is essential for reliable data interpretation, particularly with potentially degraded samples. RNAscope uses a semi-quantitative scoring system based on dots per cell rather than signal intensity [6]:
- Score 0: No staining or <1 dot/10 cells
- Score 1: 1-3 dots/cell
- Score 2: 4-9 dots/cell, few clusters
- Score 3: 10-15 dots/cell, <10% clusters
- Score 4: >15 dots/cell, >10% clusters
For degraded samples where signal may be diminished, researchers should adjust their scoring expectations accordingly and consider using digital quantification methods [5]. Advanced deep learning segmentation approaches have been developed that can outperform manual annotation, particularly for challenging cases with low-level staining [5].
Essential Research Reagent Solutions
Table: Key Reagents for RNAscope on FFPE and Degraded Samples
| Reagent/Category | Specific Examples | Function/Purpose |
|---|---|---|
| Control Probes | PPIB, POLR2A, UBC | Assess sample RNA quality and technical performance |
| Negative Controls | DapB bacterial gene | Confirm absence of background staining |
| Specialized Slides | Superfrost Plus slides | Prevent tissue detachment during multi-step procedure |
| Detection Kits | RNAscope 2.5 HD Brown/Red | Chromogenic detection for bright-field microscopy |
| Mounting Media | EcoMount, PERTEX, CytoSeal XYL | Preserve staining with appropriate refractive properties |
| Automation Systems | Ventana DISCOVERY, Leica BOND RX | Standardized processing for consistent results |
RNAscope technology represents a significant advancement for RNA biomarker detection in FFPE and degraded samples, overcoming many limitations of traditional ISH methods. The evidence demonstrates that while prolonged formalin fixation and archival storage diminish signal intensity, RNAscope remains capable of detecting RNA in tissues fixed for up to 180 days and stored for up to 15 years [27]. This capability makes it invaluable for retrospective studies utilizing archival pathology collections.
For researchers working with suboptimal samples, success depends on rigorous quality control practices, including the use of appropriate positive and negative controls, and optimization of pretreatment conditions based on sample characteristics. As the field advances, integration with digital quantification methods and deep learning approaches will further enhance the utility of RNAscope for analyzing challenging samples [5].
While RNAscope shows high concordance with PCR-based methods, its unique ability to provide spatial context for RNA expression within tissue morphology makes it a complementary technique rather than a replacement for existing methodologies. This spatial information is particularly valuable for understanding heterogeneous tissues and establishing cell-type-specific expression patterns in both research and diagnostic applications.
From Principle to Practice: Implementing RNAscope for High-Sensitivity RNA Visualization
The detection of RNA within its native tissue context is fundamental for understanding gene expression patterns, cellular heterogeneity, and disease mechanisms. For decades, traditional in situ hybridization (ISH) served as the primary method for spatial RNA detection. However, limitations in sensitivity, specificity, and reliability plagued the technique, particularly for detecting low-abundance transcripts. The advent of RNAscope technology represents a paradigm shift, addressing these core limitations through a novel probe design that enables single-molecule visualization while preserving tissue morphology. This guide provides a objective comparison of RNAscope against traditional ISH methods, framing the analysis within the broader thesis that RNAscope's standardized, one-day workflow offers superior detection sensitivity and operational efficiency crucial for modern research and drug development.
Technical Comparison: RNAscope vs. Traditional ISH
The fundamental differences between RNAscope and traditional ISH lie in their probe design and signal amplification strategies, which directly impact performance metrics such as sensitivity, specificity, and workflow robustness.
Probe Design and Signal Amplification Mechanisms
Traditional ISH relies on long, linear probes (often >500 bases) that are prone to non-specific binding and require complex optimization for each target. Its signal amplification is typically limited, leading to weak signals for low-copy RNAs.
In contrast, RNAscope utilizes a proprietary "double Z" (ZZ) probe design [30] [3] [31]. Each target is detected by a set of probe pairs that bind adjacent to each other on the target RNA. This design provides the foundation for a proprietary signal amplification system that enables simultaneous signal amplification and background suppression, allowing for single-molecule visualization [3] [32]. Each dot in an RNAscope assay represents a single RNA transcript [5].
Table 1: Core Technology Comparison
| Feature | Traditional ISH | RNAscope Technology |
|---|---|---|
| Probe Design | Long, linear probes (often >500 bases) | Short, "double Z" (ZZ) probe pairs [30] [3] |
| Signal Amplification | Limited or non-existent | Proprietary amplification via the ZZ probe binding site [3] [32] |
| Signal-to-Noise Ratio | Low, high background common | High, due to background suppression design [31] |
| Visualization | Diffuse signal | Punctate dots, each representing a single RNA molecule [5] |
Quantitative Performance Metrics
Independent studies and technical validations have consistently demonstrated RNAscope's superior performance. A key application is the precise identification of cell types, such as cardiomyocyte (CM) nuclei in cardiac regeneration studies, where antibodies against sarcomeric proteins showed a sensitivity of only 43% and specificity of 89%. Even with additional wheat germ agglutinin (WGA) staining, sensitivity only increased to 65% and specificity to 97% [7]. RNAscope intronic probes overcome these shortcomings, enabling highly specific nuclear localization [7].
Furthermore, a 2024 study in Nature Communications compared a novel multiplex method, DART-FISH, against RNAscope as a benchmark, validating RNAscope's sensitivity and specificity [33]. This establishes RNAscope as a gold standard against which newer technologies are measured.
Table 2: Performance Benchmarking
| Performance Metric | Traditional ISH | RNAscope | Supporting Data |
|---|---|---|---|
| Sensitivity | Low to moderate | Single-molecule sensitivity [31] [32] | Enables detection of low-abundance transcripts [7]. |
| Specificity | Variable, often low | High, due to ZZ probe design [3] | >70,000 probes available with a performance guarantee [34]. |
| Assay Time | Multiple days | Standardized one-day workflow | Fully automated protocols available for Leica BOND platforms [35]. |
| Multiplexing Capacity | Limited | High, with multiplex assays for simultaneous detection of multiple targets [32] | HiPlex v2 allows for detection of multiple RNA targets in a single sample [32]. |
| Probe Development | Slow, complex | Rapid (~2 weeks), with guaranteed performance [3] | Probes can be designed for almost any target in any species [3]. |
Experimental Protocols and Workflows
A direct comparison of experimental procedures highlights key differences in efficiency and standardization.
RNAscope One-Day Manual Workflow
The following protocol is adapted from standardized RNAscope procedures [7] [32] and can be completed within a single day.
Sample Preparation (Day 1, Morning)
- Tissue Fixation: Dissect tissue and fix in 4% Paraformaldehyde (PFA) for 1 hour at 4°C. For Formalin-Fixed Paraffin-Embedded (FFPE) tissues, follow standard deparaffinization and rehydration protocols.
- Cryopreparation: Saturate tissue in a sucrose series (e.g., 5%, 10%, 15%, 20% in PBS) for cryoprotection.
- Sectioning: Embed tissue in O.C.T. compound and section using a cryostat (e.g., 8-16 µm thickness). Mount sections on slides.
Pretreatment and Hybridization (Day 1, Mid-Day)
- Protease or Protease-Free Pretreatment: Apply a brief protease treatment to permeabilize the tissue. RNAscope also offers a protease-free workflow for streamlined ISH-IHC combination [32].
- Target Probe Hybridization: Apply the specific ZZ probe mix to the sections and incubate at 40°C for 2 hours.
- Signal Amplification: Perform a series of amplifier hybridizations (e.g., Amp 1, Amp 2, Amp 3) according to the specific RNAscope kit protocol. Each step typically involves a short incubation at 40°C.
Signal Detection and Counterstaining (Day 1, Afternoon)
- Chromogenic or Fluorescent Detection: For chromogenic detection (CISH), apply the enzyme-based label and chromogen (e.g., DAB). For fluorescent detection (FISH), apply fluorescently labeled probes.
- Counterstaining and Mounting: Counterstain with haematoxylin (for CISH) or a nuclear stain like DAPI (for FISH). Apply coverslips.
Protocol for Intronic RNAscope Probes for Cardiomyocyte Nuclei Identification
This specific protocol, used in cardiac regeneration studies [7], exemplifies the application of the core workflow for a challenging target.
Methodology:
- Animal Models: Wildtype or transgenic mice (e.g., FVB/NJ, Obscurin-H2B-GFP). All procedures must follow approved animal care protocols.
- Histology: Hearts are dissected, fixed in 4% PFA, saturated in a sucrose series, and frozen in O.C.T. compound. Cryosections are prepared at 16 µm thickness.
- RNAscope Assay: The standardized RNAscope fluorescent multiplex assay is performed using intronic probes designed for genes such as Tnnt2, Myl2, or Myl4.
- Microscopy and Analysis: Slides are imaged using confocal microscopy. The Tnnt2 intronic probe signal is quantified for its high colocalization with nuclear markers (e.g., Obscurin-H2B-GFP), demonstrating cardiomyocyte specificity even during mitosis.
The Scientist's Toolkit: Essential Research Reagents
Successful implementation of a standardized RNA detection workflow requires key reagents and tools. The following table details essential components for the RNAscope platform.
Table 3: Essential Research Reagent Solutions for RNAscope
| Item | Function | Example/Note |
|---|---|---|
| RNAscope Probe Sets | Target-specific ZZ probes for hybridization. | Over 70,000 unique probes available across species [34]. e.g., Tnnt2 intronic probe for CM nuclei [7]. |
| RNAscope Assay Kits | Contain amplifiers, labels, and detection reagents for signal generation. | Available as chromogenic (CISH) or fluorescent (FISH) kits, including multiplex options [32]. |
| Positive Control Probe | Verifies assay integrity and tissue RNA quality. | Probe for a ubiquitous, highly expressed gene (e.g., Polr2a). |
| Negative Control Probe | Assesses background noise and non-specific binding. | Probe for a bacterial gene (e.g., dapB) not present in the sample [5]. |
| Protease or Protease-Free Reagents | For tissue permeabilization to enable probe access. | Choice depends on sample type and whether combining with IHC [32]. |
| Hybridization Buffers | Create optimal conditions for specific probe binding. | Included in standard RNAscope kits. |
| Chromogenic or Fluorescent Substrates | For final signal visualization. | DAB (brown) for bright-field microscopy; various fluorophores for fluorescence [5] [32]. |
| Automated Staining Platform | For fully standardized, high-throughput application. | Leica BOND RX or Roche DISCOVERY ULTRA systems [35] [32]. |
Advanced Applications and Emerging Alternatives
The robustness of RNAscope has enabled its use in advanced research applications and established it as a benchmark for emerging technologies.
Key Research Applications
- Cardiac Regeneration Research: Intronic RNAscope probes (e.g., for Tnnt2) precisely identify cardiomyocyte nuclei throughout the cell cycle, overcoming the low sensitivity and specificity of antibody-based methods [7].
- Cancer Biomarker Validation: RNAscope is used in oncology to assess gene expression patterns, such as HER2 status in breast cancer, with deep learning models now being developed to automate the quantification of chromogenic RNAscope signals [5].
- Spatial Biology and Multiplexing: With the HiPlex and Fluorescent Multiplex assays, researchers can map the spatial distribution of dozens of RNA targets simultaneously within the tissue architecture, advancing the field of spatial transcriptomics [34] [32].
Comparison with Other Advanced ISH Techniques
While RNAscope is a leading solution, other advanced ISH methods have been developed.
- DART-FISH: A padlock probe-based method capable of profiling hundreds to thousands of genes in large human tissue sections. A 2024 study noted that while DART-FISH offers high multiplexing, it was validated against RNAscope, which demonstrated high sensitivity and specificity in the same study [33].
- PLISH and SABER: Other signal amplification methods reviewed by [36] as technical innovations in FISH. These methods, like RNAscope, aim to improve upon the low sensitivity of conventional ISH.
The comparative data and standardized protocols presented in this guide objectively demonstrate that RNAscope technology offers a significant advancement over traditional ISH. Its core innovation—the ZZ probe design—confers exceptional sensitivity and specificity, enabling the detection of individual RNA molecules within a morphologically preserved context. The existence of a robust, one-day workflow, supported by a vast and guaranteed probe menu and options for full automation, makes RNAscope a reliable and efficient tool. For researchers and drug development professionals requiring precise, actionable spatial gene expression data, RNAscope provides a validated and standardized solution that aligns with the demands of modern translational science.
The analysis of gene expression within its native histological context is a cornerstone of molecular pathology and biomedical research. Traditional RNA in situ hybridization (ISH) techniques, while foundational, have long been limited by significant technical challenges including insufficient sensitivity for low-abundance transcripts, high background noise from non-specific probe binding, and limited capability for multiplexing [16]. These constraints have particularly hampered the study of complex, heterogeneous tissues such as tumors and neural circuits, where understanding the co-expression patterns of multiple genes within single cells is critical for unraveling biological complexity.
The introduction of RNAscope technology in 2012 represented a paradigm shift in RNA detection methodologies [16] [2]. This novel ISH approach employs a unique probe design strategy that enables simultaneous signal amplification and background suppression, achieving single-molecule visualization while preserving tissue morphology [4]. Unlike traditional ISH methods that use single probes directly conjugated to labels, RNAscope's proprietary technology allows for highly specific multiplex detection of up to 12 RNA targets in a single sample through its HiPlex system, with 4-plex capability in its Multiplex Fluorescent v2 assay [37] [38]. This multiplexing power has transformed researchers' ability to identify cellular subpopulations, characterize co-expression patterns, and validate transcriptional profiling data within intact tissue architecture.
Technology Comparison: RNAscope Versus Alternative Approaches
Performance Metrics Across RNA Detection Methods
Table 1: Comparative analysis of RNA detection methodologies for spatial transcriptomics
| Method | Maximum Plexing | Sensitivity | Single-Cell Resolution | Tissue Compatibility | Key Limitations |
|---|---|---|---|---|---|
| Traditional ISH | Single-plex to low-plex | Low to moderate | Variable | FFPE, fresh frozen | High background, limited sensitivity, long turnaround times [16] |
| RNAscope Multiplex | 4-plex | Single-molecule detection | Excellent | FFPE, fresh frozen, cells | Requires specialized equipment and probes [37] [38] |
| RNAscope HiPlex | 12-plex | Single-molecule detection | Excellent | FFPE, fresh frozen | Longer protocol (9 hours), requires cleavable fluorophores [38] |
| BaseScope | Single-plex | Single-base resolution | Excellent | FFPE, fresh frozen | Limited to single-plexing, designed for short targets [39] |
| MERFISH/seqFISH | 100-10,000+ transcripts | Single-molecule detection | Excellent | Fixed cells/tissues | Complex barcoding, requires specialized instrumentation and computational analysis [40] |
| Live-cell Imaging | 3-5 targets | Variable | Good | Living cells | Limited multiplexing, potential perturbation of native RNA biology [40] |
Concordance with Established Gold Standard Methods
Table 2: RNAscope validation against established biomarker detection techniques
| Comparison Method | Concordance Rate | Advantages of RNAscope | Limitations/Considerations |
|---|---|---|---|
| Immunohistochemistry (IHC) | 58.7-95.3% | Direct RNA detection vs. protein; not affected by post-translational modifications or antibody quality issues [16] | Measures different biomolecules (RNA vs. protein); lower concordance expected due to biological differences in RNA-protein correlation [16] |
| qPCR/qRT-PCR | 81.8-100% | Preservation of spatial context; no RNA extraction needed; single-cell resolution [16] | Lower throughput for genome-wide studies; requires tissue sections rather than homogenized samples [16] |
| DNA ISH | 81.8-100% | Higher sensitivity for RNA detection; better signal-to-noise ratio [16] | Different applications (RNA vs. DNA detection) [16] |
The RNAscope Technology: Mechanism and Workflow
Core Principle: Dual Z-Probe Design for Unmatched Specificity
The exceptional performance of RNAscope stems from its proprietary double Z probe design, which fundamentally differs from traditional ISH approaches. Each target RNA is detected by approximately 20 pairs of "Z" probes [4] [2]. Each Z probe consists of three elements: (1) a lower region (18-25 bases) complementary to the target RNA, (2) a spacer sequence, and (3) an upper tail sequence (14 bases) [4]. The two tails from a double Z probe pair form a 28-base binding site for the pre-amplifier molecule.
This design ensures exceptional specificity through a fundamental requirement: both Z probes must bind adjacent sites on the target RNA in tandem to create a stable binding site for the pre-amplifier [4]. This mechanism dramatically reduces non-specific background because it is statistically unlikely that two independent probes will bind nonspecific targets in the correct orientation and proximity to form an amplification site. This approach is conceptually similar to fluorescence resonance energy transfer (FRET) in its requirement for dual binding events [4].
Signal Amplification Cascade for Superior Sensitivity
Following specific probe hybridization, RNAscope employs a multi-step amplification cascade that enables single-molecule detection:
- Step 1: Double Z target probes hybridize to the target RNA sequence
- Step 2: Preamplifiers hybridize to the 28-base binding site formed by each double Z probe pair
- Step 3: Multiple amplifiers bind to each preamplifier
- Step 4: Labeled probes containing fluorescent molecules or chromogenic enzymes bind to each amplifier [4]
This sequential amplification creates a dramatic signal enhancement – each target RNA molecule can bind up to 20 double Z probes, each preamplifier can bind 20 amplifiers, and each amplifier can bind 20 labeled probes, theoretically generating up to 8,000-fold signal amplification per target molecule [16]. This robust amplification enables visualization of individual RNA molecules as distinct punctate dots under a standard microscope [4].
RNAscope Signal Amplification Workflow
Experimental Workflow for Multiplex RNA Detection
The standard RNAscope multiplex fluorescent assay follows a structured workflow that can be completed within one to two days [39]:
Sample Preparation: Tissue sections (FFPE or fresh frozen) are fixed onto slides and pretreated with the RNAscope Pretreatment Kit to unmask target RNA and permeabilize cells. Protease digestion is a critical step that must be carefully optimized – under-digestion results in lower signal, while over-digestion causes poor morphology and RNA loss [15].
Probe Hybridization: Target probes (C1-C4 for 4-plex detection) are hybridized to the sample. Each target must be assigned to a specific channel with C1 serving as the mandatory primary channel. C2, C3, and C4 probes are provided as 50X concentrated stocks that must be mixed with the C1 ready-to-use probe or blank probe diluent [15].
Signal Amplification: Sequential amplification steps are performed using the RNAscope Multiplex Fluorescent Reagent Kit, with HRP blocks between channels to prevent cross-reactivity [37].
Visualization and Analysis: TSA Vivid or Opal fluorophores are used for detection, with each RNA molecule appearing as a distinct dot. Images are acquired using a fluorescent microscope with appropriate filter sets, and quantification can be performed manually or using image analysis software such as Halo or QuPath [16].
Multiplex Experimental Workflow
Essential Research Reagent Solutions
Table 3: Key reagents and materials for implementing RNAscope multiplex assays
| Reagent/Material | Function | Specific Examples | Considerations |
|---|---|---|---|
| RNAscope Multiplex Fluorescent Reagent Kit | Provides core detection reagents including pretreatment solutions, amplifiers, and HRP blocks | Catalog #323100 (20 tests) [37] | Includes TSA dilution buffer; compatible with manually assigned fluorophores |
| Target Probes | Channel-specific probes designed against target RNAs | C1 (ready-to-use), C2-C4 (50X concentrates) [37] | Must assign one target to C1 channel; species-specific design required |
| Positive Control Probes | Validate assay performance using housekeeping genes | Polr2A (low expressor), Ppib (moderate), Ubc (high) [16] | Selection should match expected expression level of targets |
| Negative Control Probes | Assess background noise using bacterial genes | Bacterial DapB gene [39] [16] | Essential for establishing specific vs. non-specific signal |
| TSA Vivid/Opal Fluorophores | Fluorescent detection of hybridized probes | TSA Vivid 520, 570, 650; Opal 520, 570, 620, 690 [37] [41] | Brightness varies; assign based on target expression level |
| Equipment | Specialized instrumentation for optimal performance | HybEZ Oven System [15] | Critical for maintaining proper temperature and humidity |
| Image Analysis Software | Quantitative analysis of punctate dots | Halo, QuPath, Aperio [16] | Enables high-throughput quantification of expression |
Strategic Implementation and Best Practices
Fluorophore Selection and Channel Assignment
Effective multiplexing requires strategic assignment of fluorophores based on their brightness and the expression levels of target RNAs. The recommended guidelines include:
- High Expression Targets: Assign to green channels (TSA Vivid 520/Opal 520), which are least distinct from tissue autofluorescence but visible to the naked eye [37]
- Medium to Low Expression Targets: Assign to orange/red channels (TSA Vivid 570/Opal 570/620) which offer good visibility with minimal background concerns [37]
- Lowest Expression Targets: Assign to far-red/near-IR channels (TSA Vivid 650/Opal 690) which are easily differentiated from autofluorescence, though not visible to the naked eye [37]
For 4-plex experiments, the RNAscope 4-Plex Ancillary Kit (Cat. No. 323120) is required in addition to the base multiplex fluorescent kit, as it contains HRP-C4 and HRP blocker necessary for the fourth channel [37].
Critical Success Factors and Troubleshooting
Several factors are crucial for achieving optimal results with RNAscope multiplex assays:
- Temperature and Humidity Control: Consistent hybridization conditions are essential. The HybEZ oven is the only hybridization system extensively validated by ACD for this purpose [15]
- Protease Optimization: Protease digestion must be carefully titrated – under-digestion reduces signal while over-digestion compromises morphology [15]
- Sample Quality: RNA integrity directly impacts results. Positive control probes (Polr2A, Ppib, Ubc) should be used to verify sample quality [16]
- Prevention of Slide Drying: Samples must not dry at any point during the procedure, and the hydrophobic barrier should remain intact throughout [15]
Integration with Immunohistochemistry
RNAscope multiplex assays can be combined with protein detection methods (ISH-IHC) for comprehensive molecular profiling [37]. This multiomic approach enables simultaneous detection of RNA and protein biomarkers within the same cellular context, providing powerful insights into gene expression regulation and protein function. The protease-free workflow option using Pretreat Pro reagent facilitates better preservation of protein epitopes while maintaining RNA detection sensitivity [41].
RNAscope technology has revolutionized multiplex RNA detection by overcoming the fundamental limitations of traditional ISH methodologies. Its proprietary double Z-probe design enables unprecedented specificity through requirement for dual probe binding, while the sequential signal amplification provides sensitivity sufficient for single-molecule detection. The technology's flexibility for 4-plex to 12-plex analysis, compatibility with standard tissue preparation methods, and ability to integrate with protein detection create a powerful platform for spatial transcriptomics.
Validation studies demonstrate high concordance with established molecular detection methods while providing the crucial advantage of spatial context preservation. For researchers investigating complex biological systems such as tumor microenvironments, neural circuits, or developmental processes, RNAscope offers the multiplexing power necessary to characterize cellular heterogeneity and interaction networks within native tissue architecture. As the field continues to advance, the technology's proven reliability and expanding capabilities position it as an essential tool for bridging the gap between bulk transcriptomic analyses and functional tissue biology.
High-throughput transcriptomic analyses, such as RNA sequencing (RNA-Seq) and microarrays, have revolutionized molecular biology by enabling researchers to study the complete set of RNA transcripts produced by the genome. These powerful techniques can generate vast amounts of data, identifying thousands of differentially expressed genes in a single experiment. However, these "grind and bind" methods have a significant limitation: they do not preserve the morphological context of the tissue, providing expression levels averaged across entire cell populations rather than at the single-cell level [42]. This averaging effect obscures critical biological information, including cellular heterogeneity, spatial distribution of expression, and specific cell-type localization of RNA molecules.
The RNAscope in situ hybridization (ISH) technology has emerged as a powerful validation tool that addresses these limitations by providing single-molecule detection within the intact tissue architecture. As a novel RNA ISH technology, RNAscope employs a unique "double Z" probe design that allows simultaneous signal amplification and background suppression to achieve single-molecule visualization while preserving tissue morphology [2]. This technical advancement enables researchers to confirm high-throughput findings at the single-cell level with spatial information, bridging the gap between bulk transcriptomic data and biological context.
This guide objectively examines the performance of RNAscope as a validation tool for RNA-Seq and microarray results, providing experimental data, detailed methodologies, and practical implementation frameworks to assist researchers in designing appropriate validation strategies for their transcriptomic studies.
Technical Comparison: RNAscope Versus Traditional Validation Methods
Performance Metrics Across Methodologies
Table 1: Comparison of RNAscope with other common gene expression analysis techniques
| Technique | Sensitivity | Spatial Context | Single-Cell Resolution | Multiplexing Capacity | Primary Applications |
|---|---|---|---|---|---|
| RNAscope | Single-molecule detection [43] | Preserved [2] | Yes [43] | Up to 12-plex with HiPlex [44] | Spatial validation, cellular localization, heterogeneity analysis |
| Traditional RNA ISH | Limited to highly expressed genes [16] | Preserved | Limited by background noise [16] | Limited | Basic localization of abundant transcripts |
| qRT-PCR | High (bulk analysis) [16] | Lost during extraction [16] | No | Limited | Bulk quantification, differential expression |
| RNA-Seq/Microarrays | Variable (technology-dependent) | Lost during processing [42] | Only with scRNA-seq | High-plex | Discovery phase, comprehensive profiling |
| IHC | Protein-level detection | Preserved | Yes | Limited by antibody availability | Protein localization, validation at protein level |
Concordance Rates with Established Techniques
Table 2: Quantitative concordance between RNAscope and established methodologies based on systematic review
| Comparison Method | Concordance Rate | Factors Influencing Concordance | Recommended Application Context |
|---|---|---|---|
| qPCR/qRT-PCR | 81.8-100% [16] | RNA quality, extraction efficiency, cellular heterogeneity | Validation of bulk expression levels |
| DNA ISH | 81.8-100% [16] | Target accessibility, probe design | Gene detection, viral identification |
| IHC | 58.7-95.3% [16] | Post-transcriptional regulation, antibody specificity | RNA-protein correlation studies |
| FISH | 97.3% in unequivocal cases [45] | Probe specificity, amplification efficiency | Clinical reflex testing, heterogeneity resolution |
The RNAscope Technology: Underlying Principles and Workflow
Molecular Mechanism of the "Double Z" Probe Design
RNAscope represents a significant advancement over traditional in situ hybridization methods through its proprietary "double Z" probe design. This innovative approach fundamentally addresses the key limitations of conventional ISH: non-specific binding and background noise. The technology employs paired "Z" probes that hybridize to adjacent sequences on the target RNA molecule. Each "Z" probe consists of three elements: (1) the lower region that hybridizes to RNA molecules, (2) a spacer (linker) sequence, and (3) the tail that binds to the pre-amplifier sequence [16].
The critical innovation lies in the requirement for dual probe hybridization before signal amplification can occur. This paired-probe system dramatically enhances specificity compared to traditional single-probe ISH approaches. The mechanism requires "Z" probes to form a dimer on the target RNA sequence before the pre-amplifier can bind and initiate the amplification cascade. This design feature makes off-target binding very unlikely and effectively suppresses background noise, enabling single-molecule detection [16] [2].
The signal amplification process contributes significantly to the technology's high sensitivity. Once bound, each RNA molecule hybridizes to 20 "Z" dimers (pre-amplifiers). Each pre-amplifier subsequently attaches to 20 amplifiers, which can then be bound by 20 labeled probes per amplifier. This cascading amplification results in up to 8,000-fold signal amplification (20 × 20 × 20 = 8,000), enabling visualization of individual RNA molecules as distinct dots under microscopy [16].
Figure 1: RNAscope "Double Z" Probe Mechanism. The diagram illustrates the sequential binding process that enables specific target detection and signal amplification.
Experimental Workflow for Validation Studies
Implementing RNAscope for validation of high-throughput data requires careful attention to workflow specifics. The process begins with slide preparation, which varies according to tissue type: formalin-fixed paraffin-embedded (FFPE) tissues (most common), tissue microarrays (TMA), fresh frozen tissues, or fixed cells [16]. Prepared slides then proceed through three critical steps: permeabilization, hybridization, and signal amplification. For larger studies, these steps can be automated using dedicated systems to ensure consistency and throughput [16].
The workflow concludes with visualization of results using either bright-field or fluorescent microscopy, depending on the probe type employed. Slides can be digitally scanned to facilitate quantification, which can be performed either manually or using specialized software applications such as Halo, QuPath, or Aperio [16] [46]. Each dot in the resulting images represents a single RNA molecule, with the number of dots correlating directly with transcript abundance [47].
Figure 2: RNAscope Experimental Workflow. The diagram outlines the key steps in implementing RNAscope for validation studies.
Implementing RNAscope for Validation of High-Throughput Findings
Experimental Design and Control Strategies
Proper experimental design is crucial for robust validation of high-throughput data using RNAscope. Each validation experiment should include three essential slide types: (1) the target marker panel, (2) a positive control, and (3) a negative control probe [47]. The positive control validates detection of a gene that should be present in the tissue, such as a housekeeping gene, and also serves as a measure of RNA integrity. The most commonly used positive controls include PPIB (for moderate expression), Polr2A (for low expression), and UBC (for high expression) [16].
The negative control probe utilizes the bacterial gene dapB (dihydrodipicolinate reductase) to confirm the absence of background noise, as this gene should not be present in animal samples [16] [47]. Proper implementation of these controls is essential for distinguishing true signal from artifacts and for verifying that experimental conditions are optimized.
When designing a validation study, researchers should select targets identified as statistically significant in high-throughput screens, prioritizing based on biological relevance and effect size. For RNA-Seq validation, typically 5-20 key targets are selected for confirmation via RNAscope, with emphasis on genes of high biological interest or unexpected expression patterns that require spatial verification.
Data Analysis and Interpretation Frameworks
RNAscope results are analyzed through quantification of labeled dots within the tissue context, with each dot representing an individual RNA molecule [47]. Analysis approaches can be categorized into two main methodologies:
Semi-quantitative histological scoring follows manufacturer-recommended criteria, where samples are scored based on the average number of dots per cell: 0 (none), 1 (1-3 dots), 2 (4-9 dots), 3 (10-15 dots), and 4 (>15 dots) [44]. This approach is particularly useful for rapid assessment and when specialized analysis software is unavailable.
Quantitative software analysis utilizes specialized platforms such as Halo, QuPath, or Aperio for precise dot enumeration and cellular segmentation [16] [46]. These tools enable more rigorous quantification, particularly valuable for heterogeneous samples or when analyzing multiple targets simultaneously.
For heterogeneous expression patterns, the Histo score (H score) can be calculated to capture both expression intensity and distribution. The H score (range 0-400) is derived using the formula: H-score = Σ (ACD score × percentage of cells per bin), providing a comprehensive metric that accounts for cellular heterogeneity [44].
Application Case Studies in Validating Transcriptomic Data
Validation of RNA-Seq Findings in Cancer Research
RNAscope has been extensively employed to validate RNA-Seq findings in various cancer types. In a notable example, researchers used RNAscope to validate results from single-cell RNA-seq analysis of spermatogonial stem cells (SSCs), confirming substantial gene expression heterogeneity among undifferentiated spermatogonia at the mRNA level and facilitating spatial localization of cell subsets [48].
In triple-negative breast cancer (TNBC), lncRNA microarrays identified over 2800 lncRNAs differentially expressed between cancerous tissue and normal adjacent tissue. Researchers employed RNAscope to confirm that expression of the lncRNA LINK-A was significantly increased in TNBC tissues compared to adjacent normal tissues, with additional spatial localization revealing expression in the cytoplasm and near the cellular membrane [42]. This spatial information significantly enhanced the biological insights gained from the microarray data alone.
Resolution of Equivocal Molecular Status in Clinical Specimens
RNAscope has demonstrated particular utility in resolving challenging diagnostic cases where traditional methods yield equivocal results. In a comprehensive study of HER2 status in invasive breast carcinoma, RNAscope was applied to quantify single-cell HER2 mRNA levels in 132 cases and compared with FDA-approved methods including FISH, IHC, and PCR [45].
The technology demonstrated 97.3% concordance with FISH in cases where FISH results were unequivocal, and proved superior to qPCR in cases with intratumoral heterogeneity or equivocal FISH results [45]. This application highlights RNAscope's potential as a reflex test within current diagnostic algorithms, particularly for cases exhibiting heterogeneity or borderline results by standard methodologies.
Viral Pathogen Detection in Infectious Disease Research
RNAscope provides a powerful approach for validating pathogen discoveries made through transcriptomic methods. In viral research, digital transcriptome subtraction (DTS) applies whole transcriptome sequencing followed by in silico removal of host sequence fragments, with remaining sequences aligned against known pathogen databases to identify candidate pathogenic sequences. RNAscope then validates the presence of these pathogenic sequences within the tissue context [49].
This approach was demonstrated in a study of high-grade gliomas, where researchers used RNAscope to validate the presence of viral pathogens identified through unmapped next-generation sequencing data [42]. The method offers both sensitivity and crucial spatial information regarding viral localization and cellular tropism.
Essential Reagents and Computational Tools
Research Reagent Solutions for RNAscope Implementation
Table 3: Essential research reagents and computational tools for RNAscope validation studies
| Item Category | Specific Examples | Function/Purpose | Implementation Notes |
|---|---|---|---|
| Control Probes | PPIB, Polr2A, UBC [16] | Positive control for RNA quality | Selection based on expected expression level |
| Negative Control | dapB bacterial gene [16] | Background assessment | Should show minimal to no staining |
| Detection Kits | RNAscope 2.5 HD BROWN/RED [44] | Chromogenic detection | Bright-field microscopy applications |
| Multiplex Kits | Multiplex Fluorescent v2 [47] | Simultaneous multi-target detection | Requires fluorescent microscopy |
| Software Tools | Halo, QuPath, Aperio [16] [46] | Image analysis and quantification | QuPath offers open-source alternative |
| Probe Design | Target-specific Z probes [2] | Gene-specific detection | Custom design for novel targets |
RNAscope technology provides a powerful methodological bridge between high-throughput transcriptomic discovery and spatial biological context. The technology's unique "double Z" probe design enables highly specific and sensitive detection of RNA targets within intact tissue architecture, addressing critical limitations of bulk analysis methods. With demonstrated applications across diverse research areas including cancer biology, infectious disease, and developmental studies, RNAscope has established itself as a robust validation platform for transcriptomic findings.
The high concordance rates with PCR-based methods (81.8-100%) support its reliability for confirming expression patterns identified through RNA-Seq and microarray analyses [16]. However, the lower concordance with IHC (58.7-95.3%) underscores the fundamental biological differences between RNA and protein detection, highlighting the complementary nature of these techniques rather than suggesting replacement [16].
For researchers implementing RNAscope validation studies, attention to proper controls, appropriate analysis methodologies, and consideration of expression heterogeneity are critical success factors. When integrated strategically within the research pipeline, RNAscope enhances the validation of high-throughput data by providing spatial context, resolving cellular heterogeneity, and ultimately strengthening biological conclusions through morphological correlation.
RNAscope represents a significant advancement in the field of in situ hybridization (ISH), addressing long-standing limitations of traditional techniques. A key challenge in biological research and diagnostics has been the robust and sensitive detection of RNA molecules within their native tissue context. Traditional RNA ISH methods, which often use digoxigenin (DIG) or radioactive probes, are hampered by high background noise and poor sensitivity, limiting their use to only highly expressed genes [16]. RNAscope, introduced by Advanced Cell Diagnostics (ACD), utilizes a novel double-Z probe design that enables simultaneous signal amplification and background suppression, allowing for single-molecule visualization while preserving tissue morphology [16] [2]. This technical overview objectively compares the performance of RNAscope against traditional alternatives, providing experimental data and methodologies that highlight its application across diverse fields including oncology, neuroscience, infectious disease, and cell & gene therapy.
Technology Comparison: RNAscope vs. Traditional ISH and IHC
The core innovation of RNAscope lies in its proprietary probe design and signal amplification system. Unlike traditional ISH, which uses a single labeled probe, RNAscope employs paired 'Z' probes that hybridize to the target RNA. This dimerization is essential for initiating a cascading amplification process, where each bound pair can ultimately recruit up to 8,000 labels, enabling the detection of individual RNA molecules as distinct dots [16]. This design inherently suppresses background noise, as off-target binding rarely results in the precise probe pairing required for amplification [2].
Table 1: Quantitative Performance Comparison of RNAscope vs. Alternative Methods
| Method | Sensitivity (Copy Number) | Specificity | Spatial Context | Multiplexing Capability | Concordance with Gold Standards |
|---|---|---|---|---|---|
| RNAscope | Single-molecule detection [16] [50] | High (probe design minimizes off-target) [2] | Preserved (full tissue context) [3] | Yes (up to 4-plex with different channels) [15] | 81.8–100% with qPCR/qRT-PCR/DNA ISH; variable with IHC [16] |
| Traditional ISH | Limited to highly expressed genes [16] | Variable (high background noise) [16] [2] | Preserved | Limited | Not systematically reported |
| IHC | Protein-level detection | Dependent on antibody quality [50] | Preserved | Yes (multiple antibodies) | 58.7–95.3% with RNAscope [16] |
| qPCR/qRT-PCR | High | High | Lost (requires tissue homogenization) [16] [3] | Not applicable | 81.8–100% with RNAscope [16] |
A systematic review comparing RNAscope to established 'gold standard' methods confirmed its high sensitivity and specificity, with a high concordance rate (81.8–100%) with qPCR, qRT-PCR, and DNA ISH [16]. Its concordance with immunohistochemistry (IHC) was lower (58.7–95.3%), which is expected as these techniques measure different biomolecules (RNA vs. protein) that may not always correlate perfectly due to post-transcriptional regulation [16]. A key advantage over IHC is that RNAscope probes can be developed for almost any target in any species within two weeks, bypassing the lengthy and often challenging process of antibody development and validation [3].
Application Spotlights and Experimental Data
Oncology
In oncology, precise biomarker detection is critical for patient stratification and therapeutic development. RNAscope enables sensitive detection of tumor-specific RNA transcripts directly in formalin-fixed paraffin-embedded (FFPE) tissue, providing valuable prognostic information.
Case Study: Validation of a DKK1 RNAscope Assay for Gastric Cancer A DKK1 RNAscope chromogenic ISH assay was developed and validated for gastric and gastroesophageal junction (G/GEJ) adenocarcinoma to identify patients for anti-DKK1 therapy [50].
- Experimental Protocol: The assay was validated on FFPE G/GEJ tumor resections according to CLIA guidelines. Specificity, sensitivity, accuracy, and precision were assessed. Tissues were fixed in 10% NBF for 16-32 hours, dehydrated, and embedded in paraffin. RNAscope was performed using a DKK1-specific probe with PPIB and dapB as positive and negative controls, respectively. Staining was quantified manually by a pathologist and digitally using QuPath software [50].
- Key Findings: The assay demonstrated a dynamic range of H-scores (0-180) in tumor cells, with signal highly specific to tumor tissue. It successfully detected cells with a single RNA molecule, confirming high sensitivity. The DKK1 RNAscope results showed a significant correlation with RNA-Seq data from the Cancer Cell Line Encyclopedia (Spearman’s rho = 0.86, p < 0.0001), supporting its accuracy [50].
- Comparative Data: RNAscope proved more sensitive than IHC, detecting DKK1 RNA in HeLa cell pellets where IHC showed no protein signal [50]. This highlights its advantage for detecting low-abundance targets.
Cell and Gene Therapy
In cell and gene therapy, characterizing differentiated cells and tracking gene expression is essential. RNAscope's ability to precisely identify cell types via specific RNA markers makes it invaluable.
Case Study: Identifying Cardiomyocyte Nuclei with Intronic Probes A major challenge in cardiac regeneration studies is the unequivocal identification of cardiomyocyte (CM) nuclei, particularly during cell cycle re-entry. Traditional antibodies to sarcomeric proteins are error-prone, and genetic models may induce unintended phenotypes [7].
- Experimental Protocol: Researchers designed intronic RNAscope probes targeting pre-mRNA of sarcomeric genes (Tnnt2, Myl2, Myl4). Embryonic and adult mouse heart cryosections were fixed in 4% PFA, saturated in sucrose, and frozen in O.C.T. compound. RNAscope was performed, and signals were visualized alongside Obscurin-H2B-GFP or cell cycle markers [7].
- Key Findings: The Tnnt2 intronic probe highly colocalized with Obscurin-H2B-GFP in adult hearts, demonstrating CM-specific nuclear labeling. In embryos, the probe labeled CM nuclei undergoing DNA replication and remained associated with CM chromatin throughout all mitotic stages, even after nuclear envelope breakdown. This "perdurance" allows reliable detection of proliferating CMs, a crucial capability for regeneration studies [7].
- Comparative Advantage: The sensitivity and specificity of intronic probes for CM nuclear identification (including during mitosis) surpasses antibody-based methods and avoids the need for genetically modified animals, presenting a powerful tool for quality control in stem cell-derived cardiomyocyte therapies [7].
Neuroscience and Infectious Disease
While the provided search results offer less direct experimental data for neuroscience and infectious disease, the established features of RNAscope make it exceptionally suitable for these fields.
- Neuroscience: The ability to perform multiplex analysis to detect several genes in a single slide is ideal for characterizing complex cell types and neuronal pathways in the brain [16] [15]. Furthermore, RNAscope's high sensitivity allows for the detection of low-abundance transcripts, such as neuropeptides and receptors, in heterogeneous neural tissues.
- Infectious Disease: RNAscope can be used to detect viral RNA in FFPE tissue specimens. Its single-molecule sensitivity enables the identification of low-level or latent infections, and its ability to provide spatial context allows researchers to pinpoint which cells are infected within a complex tissue landscape, which is crucial for understanding pathogenesis [2].
Essential Research Reagent Solutions
Successful implementation of the RNAscope assay relies on a set of specific reagents and equipment.
Table 2: Key Research Reagent Solutions for RNAscope
| Item | Function | Example/Note |
|---|---|---|
| Target Probes | Hybridize to RNA of interest | Species-specific, e.g., DKK1, Tnnt2 intronic probe [7] [50] |
| Control Probes | Assess assay performance | Positive: PPIB, POLR2A, UBC; Negative: dapB [16] [51] |
| HybEZ Oven | Maintain optimum humidity and temperature during hybridization | Critical for consistent results [51] [15] |
| Protease | Permeabilize tissue to enable probe access | Digestion time is critical; over- or under-digestion affects results [51] [15] |
| Superfrost Plus Slides | Tissue adhesion | Required to prevent tissue detachment [51] |
| ImmEdge Hydrophobic Barrier Pen | Create a barrier to retain reagents on slide | Maintains hydration; other pens may not work [51] |
| Digital Analysis Software | Quantify RNA molecules (dots/cell) | Halo, QuPath, Aperio [16] [50] [52] |
Experimental Workflow and Signaling Pathways
The standard RNAscope workflow involves critical steps from sample preparation to quantification. Adherence to the recommended protocol is essential for optimal performance.
The core technology of RNAscope can be understood through its unique probe design and the resulting amplification cascade, which is fundamental to its sensitivity.
The experimental data and comparative analysis presented in this guide demonstrate that RNAscope provides a significant technical advancement over traditional ISH and serves as a powerful complementary technique to IHC and PCR-based methods. Its high sensitivity and specificity, ability to preserve spatial information, and flexibility for multiplexing make it an indispensable tool across multiple research and diagnostic fields. In oncology, it enables precise biomarker validation; in cell and gene therapy, it allows for accurate cell identification; and in neuroscience and infectious disease, its potential for spatial transcriptomics is vast. As the technique continues to be validated in clinical settings, its role in advancing molecular pathology and therapeutic development is poised to expand further.
The integration of RNA in situ hybridization (ISH) with immunohistochemistry (IHC) represents a significant methodological advancement for researchers requiring simultaneous spatial analysis of gene expression and protein localization within intact tissue architecture. While fluorescent IHC is a well-established method for detecting protein distributions and RNAscope provides highly sensitive RNA visualization, each technique alone offers an incomplete picture of cellular states [53]. The limitations of standalone techniques become particularly evident when studying complex biological processes where transcriptional and translational regulation must be observed concurrently within specific cell types.
Traditional RNA detection methods like qPCR, while sensitive, necessitate RNA extraction and lose all spatial context [1] [16]. Conversely, conventional IHC is limited by antibody availability and specificity, with some proteins proving challenging to detect due to nonspecific antibodies or difficult-to-distinguish visualizations [53]. This gap in methodological capability is precisely where the combined RNAscope/IHC approach delivers unique value, enabling researchers to pinpoint RNA transcript locations in relation to specific protein-labeled cell types with single-molecule sensitivity [53] [16].
The RNAscope Platform
RNAscope represents a transformative advance over traditional in situ hybridization methods through its proprietary ZZ-probe design and signal amplification system. The technology employs pairs of "Z" probes that must bind adjacently to the same RNA molecule to initiate signal generation [1] [16]. Each Z probe consists of three elements: a target-specific hybridization region, a linker sequence, and a tail region that binds the amplification system [53] [16].
This dual-Z probe mechanism provides exceptional specificity because the amplification structure only binds when both probe tails are present, dramatically reducing false-positive signals from off-target binding [53] [1]. Once the Z-probe pair hybridizes to the target RNA, a multi-stage amplification cascade begins: pre-amplifiers attach to the Z-probe tails, multiple amplifiers then bind to each pre-amplifier, and finally labeled probes conjugate to the amplifiers [16]. This sophisticated process can generate up to 8,000-fold signal amplification while maintaining specificity, enabling detection of individual RNA molecules as distinct punctate dots under microscopy [16].
RNAscope Detection Capabilities by Target Length
Table 1: RNAscope Technology Variants and Their Applications
| Technology | Target Length | Probe Design | Optimal Applications |
|---|---|---|---|
| RNAscope | >300 bases (optimal: 1000 bases) | 20 ZZ pairs | Intact mRNA, long non-coding RNAs |
| BaseScope | 50-300 bases | 1-3 ZZ pairs | Short/degrated RNA, splice variants, highly homologous sequences |
| miRNAscope | ≥17 bases | Specialized design | microRNAs, small RNAs |
| DNAscope | ≥3kb (viral DNA), ≥20kb (chromosomal) | Adapted design | Viral DNA integration, chromosomal DNA |
RNAscope is specifically designed for RNA targets greater than 300 nucleotides, with optimal performance around 1000 bases using the standard 20 ZZ-pair probe design [17]. For shorter targets, ACD Bio provides specialized variants: BaseScope for sequences of 50-300 bases using 1-3 ZZ probe pairs, and miRNAscope for detecting small RNAs as short as 17 bases [17] [1]. This flexibility enables researchers to target various RNA types, from intact mRNAs to partially degraded transcripts and regulatory small RNAs.
Comparative Performance: RNAscope Versus Established Methods
Systematic Comparison with Gold Standard Techniques
A 2021 systematic review evaluating RNAscope in clinical diagnostics compared its performance against established gold standard methods including IHC, qPCR, qRT-PCR, and DNA ISH [16]. The analysis encompassed 27 retrospective studies, primarily in cancer samples, with results showing a consistently high concordance between RNAscope and molecular techniques.
Table 2: Performance Comparison Between RNAscope and Established Methods
| Comparison Method | Concordance Rate | Key Advantages of RNAscope | Limitations/Considerations |
|---|---|---|---|
| qPCR/qRT-PCR | 81.8-100% | Preserves spatial context, single-cell resolution | Different output metrics (localization vs. bulk quantification) |
| DNA ISH | High (specific rate not provided) | Superior sensitivity, single-molecule detection | - |
| IHC | 58.7-95.3% | Direct RNA detection vs. protein, different targets | Measures different biomolecules (RNA vs. protein); concordance varies based on target |
| Traditional ISH | Superior performance | Much higher sensitivity/specificity, quantitative results | Radioactive methods pose safety hazards and have poor resolution |
The review confirmed RNAscope as a highly sensitive and specific method with excellent concordance rates when compared with PCR-based techniques (81.8-100%) [16]. However, the concordance with IHC was more variable (58.7-95.3%), reflecting the fundamental difference in what each technique measures—RNA versus protein—and the complex relationship between transcript abundance and protein expression [16].
Advantages Over Traditional ISH and IHC
RNAscope addresses several critical limitations of traditional ISH, which often suffers from high background noise and limited sensitivity [1] [16]. The proprietary ZZ-probe design enables single-molecule detection while virtually eliminating off-target binding, a significant improvement over traditional approaches that use single probes directly conjugated to labels [1]. This enhanced performance comes without the safety concerns associated with radioactive ISH methods, which require special handling and offer poor spatial resolution [1].
When compared to IHC for detecting specific genes, RNAscope offers distinct advantages in situations where high-quality antibodies are unavailable or produce nonspecific staining [53]. For example, in neuroinflammation research, proteins like the inflammasome component NLRP3 and cytokine IL-1β present challenges for IHC detection due to "nonspecificity of available antibodies and the hard-to-distinguish, dot-like visualizations of the labeled proteins within the tissue" [53]. In such cases, RNAscope provides a more reliable detection method for studying expression of these important mediators.
Integrated RNAscope/IHC Protocol
Workflow for Combined Detection
The successful integration of RNAscope with IHC requires careful optimization to preserve both RNA integrity and protein antigenicity. The following workflow has been specifically validated for thicker CNS tissue sections (14μm) while minimizing tissue preparation time and costs [53].
Critical Protocol Steps and Optimizations
Tissue Preparation and Fixation: For optimal RNA and protein preservation, tissue should be fixed in fresh 10% neutral buffered formalin (NBF) for 16-32 hours at room temperature [17]. In the combined protocol for spinal cord tissue, transcardial perfusion with ice-cold 4% paraformaldehyde followed by post-fixation for 4 hours at 4°C proved effective [53]. Section thickness should be adjusted based on tissue type: 5±1μm for FFPE, 7-15μm for fixed frozen, and 10-20μm for fresh frozen tissues [17].
Protease Digestion Optimization: This represents one of the most critical steps requiring careful optimization. Under-digestion results in lower signal with ubiquitous background, while over-digestion causes poor morphology and RNA loss [15]. The combined protocol eliminates the need for RNase-removing reagents through modifications that preserve spinal cord gray matter integrity and prevent thicker sections from detaching during heat treatment [53].
Hybridization and Amplification: RNAscope procedures require the proprietary HybEZ Oven System to maintain optimal temperature (40°C) and humidity control [15] [17]. Signal amplification follows a strict sequence that must not be altered, as missing any step may result in no signal detection [15] [17]. Throughout the process, slides must not be allowed to dry completely, with a hydrophobic barrier maintained around tissues [17].
Controls and Validation: Each experiment should include three essential control slides: target marker panel, positive control (e.g., PPIB, POLR2A, or UBC based on expected expression), and negative control (bacterial dapB gene) [17] [16]. Positive control failure indicates RNA degradation, while signal in the negative control suggests background noise or nonspecific binding [16].
Experimental Demonstration: Neuroinflammation Application
Study Design and Methodology
A recently published study demonstrated the power of combined RNAscope/IHC to investigate neuroinflammatory responses in a rat model of neuropathic pain [53]. The researchers employed unilateral chronic constriction injury (CCI) of the sciatic nerve to induce neuroinflammation in the spinal cord, with tissues collected 7 days post-injury when mRNA changes are robust [53].
The experimental design capitalized on an important methodological advantage: using each animal as its own control by comparing ipsilateral (injured) and contralateral (control) sides within the same tissue section [53]. This approach guards against sampling noise across groups and requires half the costs in reagents, animal lives, and microscopy time [53].
The specific molecular targets included:
- RNA Detection: Transcripts for interleukin-1beta (IL-1β) and NLR family pyrin domain containing 3 (NLRP3)
- Protein Detection: Cell-type specific markers IBA1 (microglia) and NeuN (neurons)
Key Findings and Technical Insights
The combined approach enabled quantification of inflammatory gene expression changes within specific cell populations in somatotopically relevant laminae [53]. Researchers demonstrated that microglia are largely responsible for increased inflammatory mRNA within the spinal cord following nerve injury [53]. Furthermore, the data revealed that total increases in IL-1β and NLRP3 mRNA expression result from both increased transcription density within microglia, rather than simply from additional microglia proliferation or recruitment [53].
This application highlights particular value for targets where traditional IHC proves challenging. The authors noted that NLRP3 and IL-1β proteins are "challenging to measure accurately using immunohistochemistry due to the nonspecificity of available antibodies and the hard-to-distinguish, dot-like visualizations of the labeled proteins within the tissue" [53]. By detecting the corresponding mRNAs instead, the method provided more reliable localization and quantification of these important inflammatory mediators.
Essential Research Reagent Solutions
Table 3: Key Reagents and Equipment for RNAscope/IHC Integration
| Item | Function | Considerations |
|---|---|---|
| HybEZ II Oven System | Maintains optimal temperature/humidity | Critical for manual assays; validated specifically for RNAscope |
| RNAscope Probe Sets | Target-specific detection | Channel 1 probes are ready-to-use; C2-C4 are 50X concentrates |
| Positive Control Probes | Assay validation | PPIB (moderate expression), POLR2A (low expression), UBC (high expression) |
| Negative Control Probe (dapB) | Background assessment | Bacterial gene should not appear in animal tissues |
| SuperFrost Plus Slides | Tissue adhesion | Prevents tissue detachment, especially important for thicker sections |
| ImmEdge Hydrophobic Barrier Pen | Liquid containment | Maintains reagent coverage without drying |
| Protease Reagents | Tissue permeabilization | Requires optimization; concentration critical for signal/ morphology balance |
Visualization and Quantification Approaches
Signal Detection and Interpretation
RNAscope signals appear as punctate dots, with each dot representing a single RNA molecule [17] [16]. The critical parameter for quantification is the number of dots, not their intensity or size, which reflects the number of ZZ probes bound to each transcript rather than transcript abundance [17]. For highly expressed genes, dots may cluster together, making individual dots difficult to distinguish [16].
For multiplex assays, researchers must carefully assign probe channels based on expression levels, with higher expression genes recommended for wavelengths with potential autofluorescence (e.g., 488/green) [17]. The technology supports multiple detection methods including bright-field microscopy for chromogenic detection and epifluorescent or confocal microscopy for fluorescent applications [17].
Quantitative Analysis Methods
Analysis can be performed either manually or using specialized software platforms [16]. Manual quantification requires counting dots across multiple representative regions to obtain comprehensive results [16]. For more robust, high-throughput analysis, several software solutions are available:
- HALO (Indica Labs): Adaptable platform with powerful analytic capabilities suitable for both TMA and FFPE slides [12] [16]
- Aperio RNA ISH Algorithm (Leica Biosystems): Designed for quantitative RNA ISH analysis [12]
- QuPath: Open-source alternative for digital pathology analysis [16]
The combined RNAscope/IHC protocol enables sophisticated analyses such as quantifying RNA expression density within specific cell types defined by protein markers, providing more nuanced understanding of gene expression changes than either method alone [53].
The integration of RNAscope with IHC represents a powerful methodological advancement that enables unprecedented spatial resolution of RNA-protein relationships within intact tissues. This approach combines the exceptional sensitivity and specificity of RNAscope's ZZ-probe technology with the cell-type identification capabilities of IHC, allowing researchers to address questions about cell-type-specific gene expression that are intractable with either method alone [53].
The technology demonstrates particular value for studying low-abundance transcripts, targets with problematic antibodies, and dynamic processes like neuroinflammation where both transcriptional and translational regulation must be observed simultaneously [53]. While the method requires careful optimization of critical steps like protease digestion and stringent environmental controls, it provides researchers with a comprehensive toolkit for investigating RNA-protein colocalization with single-molecule sensitivity.
As spatial biology continues to advance, combined modality approaches like RNAscope/IHC integration will play an increasingly important role in deciphering complex biological systems, with applications ranging from basic research to drug development and clinical diagnostics.
Maximizing Signal, Minimizing Noise: A Guide to Optimizing Your RNAscope Assay
In situ hybridization (ISH) has been transformed by the development of RNAscope technology, which provides single-molecule sensitivity and single-cell resolution within the morphological context of tissue [54]. This novel ISH approach relies on a patented double Z-probe design that enables simultaneous signal amplification and background suppression, representing a significant advance over traditional RNA ISH methods [55] [6]. The critical importance of proper control implementation cannot be overstated for validating this sensitive detection system. According to technical guidelines from Advanced Cell Diagnostics (ACD), now a Bio-Techne brand, running appropriate positive and negative controls is essential for assessing sample RNA quality, optimal permeabilization, and overall assay performance [55] [6]. The control probes for PPIB (peptidylprolyl isomerase B), Polr2A (RNA polymerase II subunit A), and dapB (bacterial dihydrodipicolinate reductase) form the foundation of a robust validation system that ensures reliable interpretation of RNAscope results across diverse research applications, from viral detection to biomarker validation [49] [56] [42].
Control Probe Characteristics and Selection Criteria
Control Probe Specifications and Applications
The selection of appropriate control probes is guided by their expression characteristics and the intended experimental goals. RNAscope assays utilize a tiered system of control probes designed to validate different aspects of assay performance [55] [6].
Table 1: RNAscope Control Probe Specifications
| Control Probe | Expression Level | Copies/Cell | Primary Application | Expected Result |
|---|---|---|---|---|
| Polr2A | Low | 5-15 | Validation of low-abundance targets | Score ≥2 |
| PPIB | Medium | 10-30 | General purpose validation | Score ≥2 |
| UBC | High | >20 | Validation of high-abundance targets | Score ≥3 |
| dapB | None (bacterial) | 0 | Background assessment | Score <1 |
The negative control probe dapB targets the dihydrodipicolinate reductase gene from Bacillus subtilis strain SMY and should not generate any specific signal in properly prepared mammalian tissue samples [55] [57]. This probe serves as a critical indicator of background staining and non-specific signal, with successful assays demonstrating a dapB score of <1, indicating minimal to no background [55].
Experimental Data on Control Probe Performance
Empirical studies have validated the performance of RNAscope control probes across various tissue types and storage conditions. A 2017 study examining FFPE samples from multiple tumor types demonstrated that all three positive control probes (POLR2A, PPIB, and UBC) showed uniform expression across tissue sections, with significantly stronger signals in tumor epithelial cells compared to stromal regions [56]. Quantitative analysis revealed that for POLR2A, the lowest expressing control probe, nearly all tumor regions had at least 2 spots per cell, while PPIB expression consistently exceeded 8 spots per cell in all but a few cases [56].
Table 2: Experimental Performance of Control Probes in Tumor Tissues
| Control Probe | Tumor Region Spots/Cell | Stromal Region Spots/Cell | Expression Consistency |
|---|---|---|---|
| POLR2A | >2 (most samples) | ≥1 (most cells) | High across tumor types |
| PPIB | >8 (most samples) | >3 (variable) | Consistent through block depth |
| UBC | >15 | >6 (most samples) | Highest in tumor compartments |
This study also investigated PPIB expression at different depths within FFPE blocks (at 100-micron intervals) and found reasonably consistent expression at each level, suggesting uniform fixation throughout the tissue block [56]. Furthermore, analysis of colorectal cancer tissue microarrays containing 353 cases demonstrated that PPIB expression showed no significant degradation even in samples stored for several years, supporting the utility of RNAscope for retrospectively collected archival samples [56].
RNAscope Technology and Signaling Pathway
The Double Z-Probe Design Mechanism
The exceptional sensitivity and specificity of RNAscope technology stems from its unique double Z-probe design, which differs fundamentally from traditional ISH approaches [16]. This proprietary design utilizes paired "Z" probes that each contain three key elements: a lower region that hybridizes specifically to the target RNA sequence, a spacer linker sequence, and an upper tail region that binds to pre-amplifier molecules [16]. The critical innovation lies in the requirement that both Z-probes must bind adjacent sites on the target RNA molecule to form a stable dimer that subsequently allows pre-amplifier binding [39] [16]. This dual hybridization requirement provides the foundation for RNAscope's exceptional specificity, as off-target binding to non-specific RNA sequences rarely results in the precise pairing needed for signal amplification [58] [16].
The following diagram illustrates the RNAscope signal amplification mechanism:
Signal Amplification Cascade
Once the Z-probe dimer forms on the target RNA, a multi-stage amplification cascade begins [16]. First, the pre-amplifier molecule binds to the tail regions of the Z-probe dimer. Each pre-amplifier then provides multiple binding sites (typically 20) for amplifier molecules [39] [16]. In the final stage, each amplifier molecule accommodates numerous label probes (again, typically 20), which may be either chromogenic for bright-field microscopy or fluorescent for fluorescence detection [16]. This sequential amplification theoretically generates up to 8000-fold signal amplification per target RNA molecule (20 pre-amplifiers × 20 amplifiers × 20 label probes), enabling detection of individual RNA transcripts as distinct punctate dots [58] [16]. The entire process can be completed in a single day, a significant improvement over traditional ISH methods that often require longer hybridization and detection times [39].
Experimental Protocols for Control Implementation
Sample Preparation and Pretreatment Guidelines
Proper sample preparation is critical for successful RNAscope assays. The technology supports various sample types, including formalin-fixed paraffin-embedded (FFPE) tissues, fresh-frozen tissues, and cultured cells [55] [6]. For FFPE samples, fixation in fresh 10% neutral-buffered formalin (NBF) for 16-32 hours is recommended, followed by standard processing and embedding [6] [57]. Tissue sections should be cut at 4-5μm thickness and mounted on SuperFrost Plus slides to ensure proper adhesion throughout the rigorous assay procedure [55] [6].
Pretreatment conditions must be optimized based on tissue type and fixation quality. The standard automated protocol on the Leica BOND RX system typically includes 15 minutes of Epitope Retrieval 2 (ER2) at 95°C followed by 15 minutes of protease treatment at 40°C [55] [6]. However, adjustments may be necessary for suboptimally fixed tissues. For over-fixed tissues, increasing ER2 time in 5-minute increments and protease time in 10-minute increments is recommended while maintaining constant temperatures [55]. Conversely, for delicate tissues or under-fixed samples, a milder pretreatment of 15 minutes ER2 at 88°C with 15 minutes protease at 40°C may be appropriate [6].
Control Probe Hybridization and Detection
The RNAscope procedure involves sequential steps that must be followed precisely for optimal results [55] [6]:
Sample Pretreatment: Deparaffinization (for FFPE samples), hydrogen peroxide block, epitope retrieval, and protease digestion to permeabilize tissues and allow probe access to target RNA.
Probe Hybridization: Control probes are hybridized to target RNA for 2 hours at 40°C. For multiplex assays, channel-specific probes are mixed according to manufacturer's instructions.
Signal Amplification: A series of amplifier solutions (AMP1-AMP6 for manual assays; AMP1-AMP7 for automated systems) are applied sequentially to build the amplification cascade.
Signal Detection: Chromogenic detection uses DAB followed by hematoxylin counterstaining; fluorescent detection uses fluorophore-conjugated labels.
Mounting and Coverslipping: Appropriate mounting media must be selected based on the detection method.
Throughout the procedure, slides must not be allowed to dry out, as this causes irreversible damage and increased background [55]. The hydrophobic barrier should remain intact to maintain proper hydration, and all reagents should be fresh, particularly ethanol and xylene [6].
Research Reagent Solutions for Control Implementation
Successful implementation of RNAscope controls requires specific reagents and equipment designed to maintain assay integrity and reproducibility.
Table 3: Essential Research Reagents for RNAscope Control Assays
| Reagent/Equipment | Specification | Function in Assay |
|---|---|---|
| Control Probes | PPIB, Polr2A, UBC (species-specific), dapB | Assess RNA quality, optimize pretreatment, validate assay performance |
| Slides | Superfrost Plus | Ensure tissue adhesion throughout assay procedure |
| Hydrophobic Barrier Pen | ImmEdge Pen (Vector Laboratories) | Create barrier to maintain reagent volume and prevent drying |
| Mounting Media | Cytoseal (Brown), VectaMount PT (Red/Fluorescent) | Preserve staining and enable visualization |
| Detection System | RNAscope 2.5 HD Brown/Red or Multiplex Fluorescent | Chromogenic or fluorescent signal detection |
| Hybridization System | HybEZ Oven | Maintain optimum humidity and temperature during hybridization |
| Automation Platforms | Roche DISCOVERY ULTRA, Leica BOND RX | Standardized automated processing |
| Wash Buffer | RNAscope 1X Wash Buffer | Remove unbound reagents while maintaining tissue integrity |
Comparative Performance Data: RNAscope vs Traditional Methods
Sensitivity and Specificity Comparisons
A 2021 systematic review evaluating RNAscope in clinical diagnostics compared its performance to established "gold standard" methods including immunohistochemistry (IHC), quantitative PCR (qPCR), and traditional ISH [16]. The analysis demonstrated that RNAscope has high concordance rates with qPCR and qRT-PCR (81.8-100%) and DNA ISH (81.8-100%), confirming its reliability for nucleic acid detection [16]. However, the concordance with IHC was somewhat lower (58.7-95.3%), reflecting the fundamental difference between detecting RNA transcripts versus protein antigens [16].
The exceptional sensitivity of RNAscope enables detection of even low-abundance transcripts that may be challenging for other methods. In a study optimizing RNA-FISH for murine oocytes and embryos, researchers successfully detected and quantified low levels of Nanog mRNA in oocytes and one- and two-cell embryos, demonstrating the technique's capability for low-abundance targets [58]. The multiplex fluorescent approach allowed simultaneous detection of three housekeeping genes (Ubc, Ppib, and Polr2a) with clear differentiation of their expression levels in individual MII oocytes [58].
Advantages Over Traditional ISH and IHC
RNAscope offers several distinct advantages over traditional ISH and IHC methods [49] [16]:
Single-Molecule Sensitivity: The double Z-probe design enables detection of individual RNA molecules, far exceeding the sensitivity of traditional ISH.
Superior Specificity: The requirement for dual probe binding virtually eliminates background from non-specific hybridization.
Morphological Context: Unlike PCR-based methods, RNAscope preserves tissue architecture and enables precise cellular localization.
Multiplexing Capability: Simultaneous detection of multiple RNA targets in the same tissue section.
Species Flexibility: Probes can be developed for any species with known sequence information, overcoming antibody limitations.
For viral detection, RNAscope provides particular advantages, including the ability to differentiate between latent and active infections through sense and antisense strand detection, and identification of viral reservoirs within specific cell types [49].
Interpretation and Scoring of Control Results
Standardized Scoring Guidelines
RNAscope results are interpreted using semi-quantitative scoring guidelines that focus on counting discrete dots rather than assessing signal intensity [55] [6]. Each punctate dot represents an individual RNA molecule, with the number of dots correlating directly with transcript abundance [55]. The standardized scoring system for positive controls is as follows:
- Score 0: No staining or <1 dot per 10 cells
- Score 1: 1-3 dots/cell (visible at 20-40X magnification)
- Score 2: 4-9 dots/cell with none or very few dot clusters
- Score 3: 10-15 dots/cell with <10% dots in clusters
- Score 4: >15 dots/cell with >10% dots in clusters
For assay validation, successful PPIB staining should generate a score ≥2, while UBC should achieve a score ≥3, with relatively uniform signal distribution throughout the sample [55] [6]. The negative control dapB should yield a score of <1, indicating minimal background staining [55].
Troubleshooting Based on Control Results
Control probe results provide critical diagnostic information for troubleshooting suboptimal assays:
Low or No Signal with Positive Controls: Indicates poor RNA preservation, insufficient permeabilization, or technical issues with reagent application. Solution: Optimize protease digestion time and epitope retrieval conditions.
High Background with dapB Negative Control: Suggests inadequate washing, non-specific binding, or tissue degradation. Solution: Ensure fresh reagents, proper wash buffer preparation, and optimal fixation.
Uneven Staining Across Tissue: May result from uneven heating during hybridization or inconsistent protease digestion. Solution: Verify proper function of hybridization oven and ensure complete tissue coverage with reagents.
Proper implementation of the control system using PPIB, Polr2A, and dapB enables researchers to distinguish technical failures from true biological results, ensuring the reliability and interpretability of RNAscope data across diverse applications from basic research to clinical diagnostics.
The advent of digital image analysis (DIA) has revolutionized quantitative pathology, offering alternatives to traditional manual scoring for techniques like RNAscope. This guide objectively compares two prominent DIA platforms—the open-source QuPath and commercial HALO—against manual methods. Evidence indicates that while DIA generally outperforms manual assessment in reproducibility and throughput, platform selection depends on specific research needs. QuPath offers superior customization and cost-effectiveness, whereas HALO provides user-friendly, standardized workflows ideal for regulated environments. Within RNAscope applications, DIA enables high-throughput, single-cell resolution quantification that manual methods cannot practically achieve at scale.
Manual scoring, the traditional ground truth for histopathology quantification, is plagued by subjectivity, inter-observer variability, and labor-intensive processes, making it impractical for large-scale studies [59]. Digital image analysis mitigates these issues by providing objective, reproducible, and high-throughput quantification [60] [61]. The emergence of highly sensitive RNA in situ hybridization techniques, particularly RNAscope, which generates discrete puncta representing single RNA molecules, creates an ideal use case for DIA by providing a clear, quantifiable signal [43] [62].
This guide evaluates two distinct DIA platforms in the context of biomarker quantification and RNAscope analysis. QuPath is an open-source software celebrated for its flexibility and customizable workflows [63] [62]. In contrast, HALO is a commercial platform known for its intuitive interface and purpose-built analysis modules [63] [64]. The following sections provide a data-driven comparison to inform researchers and drug development professionals.
Table 1: Overall Software Characteristics and Performance
| Feature | QuPath | HALO | Manual Scoring |
|---|---|---|---|
| Cost | Free & Open-Source [65] | Commercial (License Fee) [65] [64] | N/A |
| Ease of Use | Steeper learning curve, requires some scripting for advanced use [65] [63] | User-friendly, minimal training required [64] | Variable, requires significant expertise |
| Flexibility & Customization | High; customizable workflows & scripts [63] [66] | Moderate; limited by available modules [63] | N/A |
| Support | Community-based forum [65] | Dedicated professional support & training [65] [64] | N/A |
| Reproducibility | High, with standardized scripts [66] | High, with standardized modules [61] | Low to Moderate [59] |
| Throughput | High, with batch processing [63] | High, with batch processing [64] | Very Low |
| Key Strength | Cost-effective, highly customizable, superior for Braak staging in neuropathology [63] | Ease of use, robust support, excels in CERAD scoring [63] | Established as historical ground truth |
Analytical Concordance and Reliability Data
Table 2: Analytical Concordance from Comparative Studies
| Measurement | QuPath vs. Manual | HALO vs. Manual | QuPath vs. HALO | Study Context |
|---|---|---|---|---|
| Cell Detection (ICC) | ρ > 0.70 [63] | ρ > 0.70 [63] | Strong correlation (ρ > 0.89) for immune cell density and spatial measures [66] | Tauopathy [63], Prostate Cancer mIF [66] |
| Ki67 Positive Cells (ICC) | N/A | N/A | N/A | Gastrointestinal NETs [59] |
| Ki67 Score (ICC) | N/A | N/A | N/A | Gastrointestinal NETs [59] |
| Agreement with Molecular Assays | N/A | Slightly better concordance & Cohen's κ with PAM50 than manual [61] | N/A | Breast Cancer Subtyping [61] |
| Prognostic Power | Essentially matched manual hazard ratios [61] | Essentially matched manual hazard ratios [61] | N/A | Breast Cancer [61] |
Experimental Protocols for Digital Image Analysis
RNAscope Image Analysis with QuPath
A standardized protocol for quantifying RNAscope-labeled neurons in rat brain using QuPath has been documented [62]. The procedure involves:
- Tissue Preparation and Staining: Fresh-frozen rodent brains are sectioned and stained using the RNAscope Fluorescent Multiplex kit. The use of negative control probes is critical for establishing analysis thresholds [62].
- Slide Scanning: Slides are digitized using a slide scanner (e.g., Zeiss AxioScan Z.1) at 20x or 40x magnification [62].
- QuPath Workflow:
- Load and Annotate: Import whole-slide images (WSIs) and annotate regions for analysis.
- Cell Detection: Use the built-in cell detection algorithm. Key parameters include cell expansion, background radius, and intensity thresholds, which must be optimized and validated for each tissue and probe [62].
- Set Positivity Threshold: The fluorescence intensity threshold for classifying a cell as "positive" is derived from negative control slides. This standardizes quantification and improves reproducibility [62].
- Run Analysis and Export Data: Execute the analysis and export cell-by-cell data, including transcript counts and cell coordinates, for further statistical analysis.
HALO Analysis for Multiplex Fluorescence
HALO utilizes a module-based approach, such as the Highplex FL or FISH-IF module for multiplex RNAscope analysis [64]. The typical workflow is:
- Image Import and Quality Check: Load WSIs and use channel visualization tools to assess image quality [64].
- Select Analysis Module: Choose the appropriate, pre-configured module.
- Segment Cells: Use pre-trained AI networks within the module to optimize nuclear and/or cytoplasmic segmentation. The software provides real-time tuning for parameter adjustment [64].
- Define Phenotypes: Use the phenotype editor to assign cell types based on marker combinations using an intuitive grid interface [64].
- Run Analysis and Review: Execute the analysis. Results can be interactively explored via markup images that link cell data back to its spatial context, allowing for visual validation and data mining [64].
Protocol for Comparative Validation (QuPath vs. HALO)
A recent neuropathology study established a robust protocol for comparing QuPath and HALO [63]:
- Tissue and Staining: 70 slides from 35 cases (hippocampus and mid-frontal gyrus) were stained for AT8 (tau) and digitized at 20x magnification.
- Analysis Alignment: Both platforms were used to calculate percent positivity, object density, and optical density on the same tissue regions.
- Validation: Results from both software were validated against manual counts and correlated with established neuropathological scores (Braak stage and CERAD score). Percent positivity was found to be the most consistent and reliable metric across platforms [63].
Visualization of Workflows and Relationships
RNAscope DIA Quantification Workflow
Diagram 1: RNAscope DIA Workflow. This diagram outlines the core steps for quantifying RNAscope assays using either QuPath or HALO, highlighting the step of using negative controls to set thresholds.
Decision Logic for Platform Selection
Diagram 2: Platform Selection Logic. A decision tree to guide researchers in choosing between QuPath and HALO based on budget, technical resources, and analytical needs.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Key Reagents and Materials for RNAscope and DIA
| Item | Function / Application | Example Product / Specification |
|---|---|---|
| RNAscope Fluorescent Multiplex Kit | Core reagent kit for sensitive, multiplex RNA in situ hybridization. | RNAscope Fluorescent Multiplex Reagent Kit (ACD) [62] |
| Target Probes | Probes bind specific RNA targets; each is assigned a channel (C1, C2, C3). | RNAscope Target Probes (e.g., Rn-Hcrtr1-C1) [62] |
| Negative Control Probe | Essential for determining background signal and setting positivity thresholds in DIA. | RNAscope 3-plex Negative Control Probe [62] |
| Positive Control Probe | Verifies assay integrity and tissue RNA quality. | Species-specific positive control probes (e.g., Ppib) |
| HybEZ Oven | Specialized hybridization oven for standardized and optimized RNAscope assay conditions. | HybEZ II System (ACD) [62] |
| High-Performance Microscope Slide | Ensures tissue adhesion and minimizes background during stringent assay steps. | Fisherbrand Superfrost Plus Microscope Slides [62] |
| Aqueous Mounting Medium with DAPI | Preserves fluorescence and counterstains nuclei for cell segmentation in DIA. | Fluoro-Gel II with DAPI [62] |
| Slide Scanner | Creates high-resolution digital whole-slide images for software analysis. | Zeiss AxioScan Z.1, Hamamatsu Nanozoomer, etc. [62] |
| Computer Workstation | Runs DIA software; requires substantial RAM and processing power for large images. | 64-bit OS, 16+ GB RAM, fast multicore processor [62] |
The transition from manual scoring to digital image analysis represents a significant advancement for pathological quantification, particularly for precise techniques like RNAscope. Both QuPath and HALO are powerful platforms that deliver superior objectivity, reproducibility, and efficiency compared to manual methods.
The choice between them is not a matter of absolute superiority but of strategic fit. QuPath is the optimal choice for resource-limited environments, advanced users requiring deep customization, and projects involving large-scale or novel analytical workflows. HALO justifies its cost for labs requiring a standardized, user-friendly platform with robust commercial support, especially in regulated or multi-user environments. Researchers must weigh their specific needs regarding budget, technical expertise, and analytical requirements to select the most effective tool for their research.
Thesis Context: RNAscope technology represents a major advance over traditional in situ hybridization (ISH), offering unparalleled sensitivity and specificity for RNA visualization in intact cells and tissues. This guide explores how its flexible probe design strategy empowers this performance and provides a framework for selecting between its catalog and custom-made probe options.
Table of Contents
- RNAscope: A Revolution in Signal-to-Noise Ratio
- The Two Paths: Catalog Probes vs. Made-to-Order Probes
- Comparative Performance Data: Catalog vs. Custom Probes
- Experimental Protocols for Probe Validation
- Research Reagent Solutions
- Decision Framework and Conclusion
RNAscope: A Revolution in Signal-to-Noise Ratio
Traditional RNA ISH methods, which use single probes labeled with digoxigenin or radioisotopes, are often plagued by high background noise and poor sensitivity, limiting their reliability for detecting low-abundance transcripts [16]. RNAscope overcomes these limitations through a proprietary double Z probe design that fundamentally improves the signal-to-noise ratio [4].
This patented method uses probe pairs that must bind in tandem to the target RNA to initiate a signal amplification cascade. It is highly improbable for two independent probes to bind non-specifically at adjacent sites, which suppresses background noise. Once bound, the system can amplify the signal up to 8,000 times, enabling the visualization of single RNA molecules as distinct, punctate dots under a microscope [16] [4]. This core technological advance makes RNAscope both highly sensitive and specific, forming the foundation upon which all RNAscope probes are built.
The Two Paths: Catalog Probes vs. Made-to-Order Probes
ACD's probe ecosystem is divided into two main categories, summarized in the table below, which serve complementary needs in the research pipeline.
Table 1: Comparison of RNAscope Catalog and Made-to-Order Probes
| Feature | Catalog Probes | Made-to-Order Probes |
|---|---|---|
| Definition | Pre-designed, validated probes for well-established targets [67] | Newly designed probes for a researcher's specific gene of interest [67] |
| Key Advantage | Speed and convenience; ship in ~2 weeks [67] | Flexibility; enables research on any gene from any species [67] [19] |
| Turnaround Time | Approximately 2 weeks [67] | Approximately 2 weeks for design and manufacture [67] [49] |
| Scope | >40,000 RNA targets across >400 species and >100 viruses [67] | Virtually any gene in any genome for visualization in any tissue or cells [67] |
| Ideal Use Case | Validating known biomarkers, routine assays, and initial method development | Novel target discovery, studying splice variants, proprietary genes, or uncommon species |
The "Z" probe design is the cornerstone of both probe types. The following diagram illustrates how this unique architecture enables specific signal amplification.
Comparative Performance Data: Catalog vs. Custom Probes
A systematic review evaluated RNAscope's performance against gold-standard techniques like IHC, qPCR, and DNA ISH. The review, which included studies using both catalog and the underlying custom probe design pipeline, confirmed RNAscope's high reliability [16].
Table 2: RNAscope Concordance with Gold-Standard Techniques
| Technique | Concordance Rate (CR) with RNAscope | Primary Reason for Discrepancy |
|---|---|---|
| qPCR / qRT-PCR | 81.8% - 100% [16] | Different measured analytes (RNA vs. extracted RNA) and platform sensitivities. |
| DNA ISH | 81.8% - 100% [16] | High specificity of both in-situ techniques. |
| IHC | 58.7% - 95.3% [16] | Different measured analytes (RNA vs. protein); discordance can reveal post-transcriptional regulation. |
A powerful example of a custom-made probe is the intronic probe. Researchers designed custom RNAscope probes targeting intronic regions of genes like Tnnt2 (cardiac troponin T) to specifically label cardiomyocyte nuclei. This approach bypassed the limitations of antibody-based nuclear labeling, achieving high specificity and sensitivity for identifying cycling cardiomyocytes, a task that was previously technically challenging [23].
Experimental Protocols for Probe Validation
Whether using a catalog or custom probe, a rigorous experimental protocol is essential for generating reliable data. The following workflow, applicable to both probe types, is adapted from established methods for multiplexed RNAscope [68] [23].
Protocol Workflow:
Detailed Methodology
Tissue Preparation:
- For formalin-fixed paraffin-embedded (FFPE) tissues, section at 5 µm. For frozen tissues, section at 10-20 µm [23].
- Fix tissues in 4% paraformaldehyde (PFA) for 1 hour at room temperature or overnight at 4°C [23].
- Perform slide pretreatment with RNAscope Pretreatment Kit to unmask target RNA and permeabilize cells [4].
Probe Hybridization:
Signal Amplification:
Visualization and Quantification:
Key Experimental Controls
- Positive Control Probe: A probe for a ubiquitous housekeeping gene like PPIB validates assay success and tissue RNA integrity [16].
- Negative Control Probe: A probe for a bacterial gene like dapB confirms the absence of non-specific background staining [16].
Research Reagent Solutions
The following table details essential materials and their functions for performing an RNAscope experiment.
Table 3: Essential Research Reagent Solutions for RNAscope
| Item | Function / Description | Example Products / Targets |
|---|---|---|
| RNAscope Probe | The core reagent that specifically hybridizes to the target RNA. | Catalog: Hs-MKI67 (Human Ki-67). Custom: Tnnt2 intronic probe [67] [23]. |
| Detection Kit | Contains amplifiers and labels for signal generation. | RNAscope 2.5 HD Reagent Kit (BROWN), Multiplex Fluorescent Reagent Kit v2 [67]. |
| Control Probes | Validate assay performance and specificity. | PPIB (Positive Control), dapB (Negative Control) [16]. |
| Pretreatment Kit | Unmasks target RNA and permeabilizes cells for probe access. | RNAscope Pretreatment Kit [4]. |
| Image Analysis Software | Enables precise, quantitative analysis of RNA signals. | HALO, QuPath, Aperio [16]. |
Choosing between a catalog and a made-to-order probe depends on your project's specific needs. The following decision framework can help guide your selection.
Conclusion: The choice between catalog and made-to-order RNAscope probes is not a matter of quality but of application. Catalog probes offer a validated, off-the-shelf solution for studying established targets, maximizing efficiency. Made-to-order probes provide unparalleled flexibility to explore novel genes, splice variants, and proprietary targets, pushing the boundaries of discovery. Both options are powered by the same robust, sensitive, and specific RNAscope technology that has definitively surpassed the capabilities of traditional ISH. By aligning your probe choice with your experimental goals, you can fully leverage this powerful spatial genomics platform.
This guide provides an objective comparison of RNAscope in situ hybridization (ISH) technology against traditional ISH methods, focusing on the critical balance between financial and time investments and the resulting quality of gene expression data. The analysis confirms that while RNAscope requires a significant initial investment in specialized probes and optimization, its superior sensitivity and specificity lead to more reliable data, potentially reducing long-term costs associated with experimental repetition and erroneous conclusions. For research and drug development requiring precise spatial gene expression analysis at single-cell resolution, RNAscope presents a compelling value proposition that justifies its costs.
In the fields of cancer research, neuroscience, and drug development, accurate visualization of gene expression within its native tissue context is paramount. For decades, traditional RNA in situ hybridization (ISH) has been a fundamental technique but has been hampered by significant limitations in sensitivity and specificity, often leading to high background noise and an inability to detect low-abundance transcripts [16]. These technical challenges directly impact data quality, potentially compromising target validation, biomarker discovery, and therapeutic development.
The advent of RNAscope technology (Advanced Cell Diagnostics, a Bio-Techne brand) introduced a novel, highly sensitive approach to RNA ISH. However, researchers must weigh its enhanced performance against greater monetary and time investments. This analysis objectively compares RNAscope with traditional ISH methods, evaluating costs in the context of the resulting data quality—a critical consideration for researchers and drug development professionals allocating limited resources.
Technology Comparison: RNAscope vs. Traditional ISH
Fundamental Technological Differences
The core difference lies in probe design and signal amplification. Traditional ISH often uses single, directly labeled probes, which are prone to non-specific binding and offer limited signal amplification. In contrast, RNAscope employs a proprietary "double Z" probe design, which requires two independent probes to bind in tandem to the target RNA before a powerful signal amplification cascade can proceed [16] [30]. This fundamental distinction is the primary driver of differences in performance, cost, and workflow.
RNAscope Signaling Pathway
The following diagram illustrates the key steps in the RNAscope signal amplification process, which is central to its high sensitivity and specificity.
The following table summarizes key performance metrics based on published comparative studies and technical data.
Table 1: Quantitative Performance Comparison of RNA ISH Techniques
| Performance Parameter | Traditional ISH | RNAscope | Experimental Basis |
|---|---|---|---|
| Analytical Sensitivity | Low; limited to highly expressed genes [16] | High; capable of single-molecule detection [16] [30] | Systematic review; manufacturer validation |
| Specificity | Variable; high non-specific background common [16] | High; nearly 100% due to dual Z-probe design [16] | Systematic review comparing to IHC, qPCR |
| Concordance with qPCR/qRT-PCR | Not systematically reported | 81.8% - 100% [16] | Narratively reviewed concordance rates |
| Concordance with IHC | Not systematically reported | 58.7% - 95.3% [16] (Note: measures RNA vs. protein) | Narratively reviewed concordance rates |
| Single-Cell Resolution | Limited | Excellent; allows quantification of heterogeneity [16] [69] | Cell type-specific quantification frameworks (e.g., QuantISH) |
| Applicability to FFPE Tissues | Challenging due to RNA degradation | Robust; works well on FFPE archives [16] [70] | Widespread use in retrospective FFPE studies |
Experimental Protocols for Performance Validation
Core RNAscope Workflow Protocol
The standard RNAscope assay for Formalin-Fixed Paraffin-Embedded (FFPE) tissues involves a single-day workflow, similar to immunohistochemistry (IHC) [16] [29]. The critical steps where protocol differs significantly from traditional ISH are outlined below.
RNAscope Experimental Workflow
- Slide Preparation and Pretreatment: FFPE sections are baked, deparaffinized, and rehydrated. A critical, often optimized step follows: pretreatment with target retrieval reagents and proteases to expose target RNA sequences. The necessity for optimization here is a key time cost [29].
- Probe Hybridization: Target-specific, custom-designed "Z" probe pairs are hybridized to the RNA for 2 hours at 40°C [16].
- Signal Amplification: A series of amplifier molecules are hybridized sequentially to the bound probe tails. This multistep process, while time-consuming, provides up to 8,000-fold signal amplification [16].
- Detection and Counterstaining: Signals are developed using chromogenic (e.g., Fast Red) or fluorescent labels, followed by counterstaining (e.g., hematoxylin) [71].
- Analysis: Each dot represents a single RNA molecule. Quantification can be manual or using specialized software (e.g., Halo, QuPath, Aperio) for high-throughput, reproducible results [16] [69].
Critical Quality Control Experiments
A rigorous quality control protocol is non-negotiable for ensuring data quality and is a fundamental part of the RNAscope workflow [29].
Table 2: Essential Quality Control Measures for RNAscope
| QC Component | Purpose | Recommended Protocol & Interpretation |
|---|---|---|
| Positive Control Probe | Validate assay technique and sample RNA quality. | Use housekeeping gene probes (e.g., PPIB for moderate expression, Polr2A for low expression). Strong staining indicates success; failure suggests RNA degradation or technical error [16] [29]. |
| Negative Control Probe | Assess background noise and non-specific binding. | Use a bacterial gene probe (e.g., DapB). The absence of staining confirms assay specificity [16] [29]. |
| Pretreatment Optimization | Balance RNA exposure with tissue preservation. | Titrate protease digestion time using control probes. Aim for high positive control signal with no negative control background [29]. |
Cost-Benefit Analysis: Investment vs. Data Quality
Monetary and Time Investments
The financial and temporal costs of RNAscope are substantial and must be factored into project planning.
Direct Monetary Costs:
- Probes: Custom target-specific probes represent a significant recurring cost. RNAscope probes are more complex and expensive than those for traditional ISH.
- Reagent Kits: The specialized amplification kits are a major cost component.
- Specialized Equipment: While standard bright-field or fluorescent microscopes suffice, automated staining systems (e.g., from Leica or Roche) and slide scanners add capital expenditure but improve reproducibility [72].
- Analysis Software: Licenses for advanced image analysis platforms (e.g., Halo, Indica Labs) add to the cost but enable robust quantification [69].
Time Investments:
- Assay Development: Protocol optimization, especially pretreatment conditions for new tissue types, can take days to weeks [29] [71].
- Probe Validation: For custom targets, validating probe functionality is a critical, time-consuming step. Novel protocols using cell-free synthesized mRNA can accelerate this process [71].
- Assay Runtime: The core hybridization and amplification protocol is typically completed in one day, comparable to IHC [16].
- Data Analysis: Manual dot counting is laborious. Automated analysis requires initial setup time but drastically reduces analysis time thereafter [69].
Benefits and Data Quality Returns
The investment in RNAscope is offset by clear benefits that directly impact research outcomes and decision-making.
- Superior Data Quality: The high sensitivity and specificity minimize false negatives and positives, leading to more trustworthy data [16].
- Reduced Long-Term Costs: Reliable results reduce the need for costly reagent repeats and prevent misguided research directions based on poor-quality data. One systematic review suggests it could complement or confirm unclear results from IHC or qPCR, potentially saving resources [16].
- Access to Previously Undetectable Targets: The ability to detect low-abundance transcripts, short sequences, and splice variants opens new research avenues that are inaccessible with traditional ISH [71].
- Quantitative and Spatial Data: The single-molecule resolution allows for true quantification of gene expression within specific cell types in the tissue context, providing insights into tumor heterogeneity [69].
The Scientist's Toolkit: Essential Research Reagent Solutions
Table 3: Key Reagents and Materials for RNAscope Experiments
| Item | Function/Benefit | Considerations for Cost & Quality |
|---|---|---|
| RNAscope Assay Kits | Provide all necessary reagents for the hybridization and amplification steps in a standardized format. | Essential for consistency. Bulk purchasing can reduce cost per sample. |
| Target-Specific Probes | Core reagent that defines the target mRNA sequence for detection. | The major cost driver. Validated probes for common targets are available; custom designs are more expensive. |
| Control Probes (PPIB, Polr2A, DapB) | Critical for validating experimental conditions, sample quality, and assay specificity [29]. | Non-negotiable for rigorous science. Failure to use controls risks generating unreliable data. |
| Optimized Tissues (e.g., Tissue Microarrays) | Pre-validated tissues with known RNA quality [70]. | Higher initial cost but saves time and resources by guaranteeing sample quality and eliminating optimization steps. |
| Automated Image Analysis Software (e.g., Halo, QuPath) | Enables high-throughput, objective quantification of RNA signals, reducing analyst bias and time [16] [69]. | License fees are a cost, but they pay for themselves in saved personnel time and improved reproducibility. |
For research and diagnostic applications where precise spatial localization and accurate quantification of RNA are critical, RNAscope technology offers a significant advantage in data quality over traditional ISH. The initial and recurring costs are higher, and the protocol requires careful optimization. However, the return on investment is realized through highly reliable, publication-quality data, the ability to study low-expression targets and heterogeneity, and potential long-term savings from fewer experimental repeats.
The ongoing development of more automated workflows [70], advanced analysis pipelines like QuantISH [69], and expansion into the clinical diagnostics sphere [16] [72] promise to further enhance the value proposition of RNAscope, solidifying its role as a powerful tool in modern molecular pathology and life science research.
For researchers investigating spatial gene expression, RNA in situ hybridization (ISH) has long been a fundamental technique, yet one plagued by inherent technical challenges. The central dilemma in traditional ISH methodologies involves balancing the competing demands of detection sensitivity and morphological preservation while minimizing background noise. While techniques like immunohistochemistry (IHC) provide morphological context, they rely on antibody availability and may lack specificity, particularly for species beyond human, rat, and mouse [3]. Similarly, molecular profiling methods like PCR and microarrays, while providing quantitative data, sacrifice crucial spatial context and information about expression heterogeneity [3]. The emergence of the RNAscope assay represents a technological paradigm shift, introducing a proprietary probe design that simultaneously amplifies target-specific signals while systematically suppressing background noise from non-specific hybridization [32]. This guide objectively compares the performance of the RNAscope platform against traditional ISH and alternative detection methods, providing supporting experimental data and detailed protocols to frame its advantages within the broader thesis of achieving superior detection sensitivity without compromising tissue integrity.
The Fundamental Challenge: Technical Limitations of Traditional ISH
Traditional ISH techniques have been limited by several interconnected pitfalls that directly impact data reliability and interpretation. A primary concern is high background noise stemming from non-specific probe hybridization, which can obscure genuine signals and lead to false positives [3]. This is compounded by inadequate sensitivity for detecting low-abundance transcripts, a significant limitation when studying subtle regulatory changes or heterogeneous cell populations [39]. Furthermore, many conventional approaches suffer from lengthy turnaround times and are restricted to single-gene analysis per section, inefficiently utilizing precious tissue samples [39].
The problem of accurately attributing signals to specific cell types is starkly illustrated in cardiac regeneration research. One study estimated that using antibodies to sarcomeric proteins to identify cardiomyocyte nuclei achieved a sensitivity of only 43% and specificity of 89%, figures that improved only to 65% and 97%, respectively, even when combined with wheat germ agglutinin (WGA) staining to outline cell membranes [7]. This level of uncertainty becomes critically problematic when attempting to identify rare events like cell cycle re-entry in adult cardiomyocytes.
The RNAscope Solution: A Paradigm Shift in Probe Design and Signal Amplification
The RNAscope platform addresses fundamental limitations of traditional ISH through its patented double-Z ("ZZ") probe design [39] [32]. This technology employs pairs of oligonucleotides (18-25 bases each) that are complementary to approximately 50 contiguous bases in the target RNA [39]. The core innovation lies in the requirement for both probes in a pair to bind adjacent to each other on the target RNA before signal amplification can proceed. This dual-binding mechanism provides the foundation for exceptional specificity, as off-target hybridization to non-specific RNA sequences does not result in signal amplification [39].
The sequential amplification process theoretically yields an 8000-fold increase in signal per target, enabling detection of single transcripts with single-molecule sensitivity [39] [3]. Unlike traditional ISH methods where background suppression often compromises signal strength, RNAscope's design decouples these two parameters, allowing for simultaneous high signal amplification and low background noise. The platform's flexibility supports detection of multiple RNA targets in a single assay (multiplexing) while preserving tissue morphology across various sample types, including fresh-frozen and formalin-fixed paraffin-embedded (FFPE) tissues [39] [32].
Table 1: Key Advantages of RNAscope Over Traditional ISH Methods
| Feature | Traditional ISH | RNAscope Technology |
|---|---|---|
| Probe Design | Single oligonucleotides or cRNAs | Patented double-Z ("ZZ") probe pairs [39] |
| Specificity Mechanism | Variable; often high background | Requires dual probe binding; minimal non-specific amplification [39] |
| Sensitivity | Limited for low-abundance targets | Single-molecule detection [39] [3] |
| Multiplexing Capacity | Typically single-plex | Simultaneous detection of up to 12 targets [32] |
| Turnaround Time | Often multiple days | One-day protocol [39] |
| Morphology Preservation | Variable, often compromised | Excellent across tissue types [3] [32] |
Diagram 1: RNAscope Signal Amplification Pathway. The proprietary double-Z probe design requires two probes to bind adjacent target sequences before sequential amplification can occur, ensuring high specificity [39] [32].
Performance Comparison: Quantitative Data and Experimental Evidence
Enhanced Specificity in Challenging Applications
The superior performance of RNAscope is particularly evident in applications where specificity is paramount. In cardiac regeneration studies, researchers developed intronic RNAscope probes targeting Tnnt2, Myl2, and Myl4 to precisely identify cardiomyocyte nuclei—a task notoriously challenging with antibody-based methods. The Tnnt2 intronic probe demonstrated high colocalization with Obscurin-H2B-GFP in adult mouse hearts, confirming cardiomyocyte specificity [7]. Crucially, these intronic probes remained associated with cardiomyocyte chromatin throughout all mitotic stages, including after nuclear envelope breakdown, enabling reliable investigation of DNA synthesis and mitotic activity even in challenging post-infarction environments with high background autofluorescence [7].
Correlation with qRT-PCR and Quantification Capabilities
The quantification capabilities of RNAscope were rigorously evaluated in a study comparing it with qRT-PCR for analyzing colorectal cancer biomarkers in archival FFPE tissue. The research demonstrated that multiple image analysis tools could reliably quantify RNAscope signals, with the WEKA tool showing the greatest agreement with manual quantification [73]. Most significantly, when quantifying expression of MLH1, GFI1, and TNFRSF11A in colorectal cell lines, the image analysis methods applied to RNAscope data performed at a similar level to qRT-PCR, establishing its credibility for quantitative expression analysis while preserving spatial information that PCR-based methods destroy [73].
Table 2: Performance Comparison Across RNA Detection Platforms
| Parameter | qRT-PCR | Traditional ISH | RNAscope |
|---|---|---|---|
| Sensitivity | High (detects low abundance) | Low to Moderate | High (single-molecule) [39] |
| Spatial Context | No preservation | Preservation with variable quality | Excellent preservation [73] |
| Quantification | Excellent | Limited | Good correlation with qRT-PCR [73] |
| Specificity | High | Variable | High (double-Z probe design) [39] |
| Multiplexing | Limited (with controls) | Typically single-plex | Up to 12-plex [32] |
| Tissue Requirements | Homogenized (context lost) | Intact tissue | Intact tissue (FFPE/frozen) [39] [32] |
Sample Preparation and Protocol Optimization: Preserving Morphology and RNA Integrity
Tissue Preservation Considerations
Optimal RNAscope results begin with proper tissue preservation. While FFPE tissues are widely used, recent comparative studies have revealed that alternative preservatives may offer advantages for molecular integrity. Research comparing 10% neutral buffered formalin (NBF) to silver nanoparticles (AgNPs) demonstrated that AgNPs solution maintained consistent DNA, RNA, and protein concentrations across heart, liver, and kidney tissues over 72 hours, whereas formalin treatment led to significant degradation over time [74]. However, 10% NBF demonstrated superior preservation of tissue morphology, highlighting the trade-offs between molecular and structural preservation [74].
Detailed Protocol for Fresh-Frozen Tissues
The following protocol, validated for fresh-frozen mouse brain tissue, can be adapted for other tissues with appropriate optimization [75]:
Tissue Preparation and Sectioning:
- Perfuse mouse with 1x PBS and harvest tissue immediately.
- Embed tissue in OCT compound and freeze in 2-methylbutane surrounded by dry ice in methanol.
- Store at -80°C overnight.
- Section on a cryostat at 16 μm thickness onto SuperFrost Plus slides.
- Store slides at -80°C until use.
Sample Fixation and Dehydration:
- Immerse slides in 4% PFA in 1x PBS for 2 hours at room temperature.
- Wash twice in 1x PBS.
- Dehydrate through graded ethanols: 50% EtOH (5 min), 70% EtOH (5 min), and two changes of 100% EtOH (5 min each).
- If necessary, store slides in 100% EtOH at -20°C for up to one week (not recommended for optimal results).
Tissue Pretreatment:
- Apply RNAscope Hydrogen Peroxide for 10 minutes at room temperature to block endogenous peroxidase activity.
- Wash twice in distilled water for 2 minutes each.
- Apply Protease Plus for 10 minutes at room temperature to permeabilize tissues.
- Wash twice in 1x PBS for 2 minutes each.
Hybridization and Signal Amplification:
- Prepare probe master mix (C1:C2:C3 ratio of 50:1:1 for multiplexing).
- Apply probe mix to slides and incubate for 2 hours at 40°C in a HybEZ oven.
- Wash twice in 1x wash buffer for 2 minutes each.
- Perform sequential amplifications: AMP1 (30 min, 40°C), AMP2 (30 min, 40°C), AMP3 (15 min, 40°C), with washes between steps.
- Develop HRP signals using fluorophore-conjugated tyramides (e.g., Opal dyes diluted 1:750 in TSA buffer).
- Counterstain with DAPI (30 seconds to 5 minutes) and mount with ProLong Gold antifade reagent.
Diagram 2: RNAscope Workflow for Fresh-Frozen Tissues. This optimized protocol maintains RNA integrity and tissue morphology through controlled fixation and dehydration steps [75].
The Scientist's Toolkit: Essential Research Reagent Solutions
Successful implementation of RNAscope technology requires specific reagents and equipment designed to optimize performance and maintain experimental consistency.
Table 3: Essential Research Reagents for RNAscope Implementation
| Reagent/Equipment | Function | Application Notes |
|---|---|---|
| RNAscope Multiplex Fluorescent Kit | Core reagents for detection | Contains AMPs, HRPs, and blockers for 3-plex detection [75] |
| Target Probes | Gene-specific detection | Designed for C1, C2, C3 channels; C1 most sensitive [39] |
| Protease Plus | Tissue permeabilization | Critical for probe accessibility; overtreatment damages morphology [75] |
| Hydrogen Peroxide | Endogenous peroxidase block | Reduces background in fluorescent detection [76] |
| HybEZ Oven | Controlled hybridization | Maintains precise 40°C with humidity control [75] |
| Hydrophobic Barrier Pen | Defines hybridization area | Prevents reagent evaporation and cross-contamination [75] |
| Opal Fluorophores | Signal detection | Used with HRC-C1/C2/C3 at 1:750 dilution in TSA buffer [75] |
| Positive/Negative Control Probes | Assay validation | Essential for establishing protocol effectiveness [39] |
Advanced Applications and Future Directions
The adaptability of RNAscope continues to expand its applications across research domains. The development of BaseScope, a newer ultrasensitive platform, allows discrimination of single nucleotide polymorphisms or splice variants that differ by short exons through improved amplification chemistry of single oligonucleotide probe pairs [39]. While currently limited to single-plex analysis, this technology enables investigation of previously undetectable RNAs, including non-coding RNAs and specific splice variants [39].
In tissue clearing and 3D imaging applications, emerging methods like OptiMuS-prime show promise for compatibility with RNAscope. This novel passive tissue-clearing technique replaces SDS with sodium cholate (SC) combined with urea, achieving better reagent infiltration while retaining structural integrity and preserving RNA for detection [77]. Such advancements highlight the ongoing convergence of spatial transcriptomics with volumetric imaging, opening new possibilities for comprehensive tissue analysis.
The RNAscope platform represents a significant advancement in RNA ISH technology, effectively addressing the common pitfalls of background noise and morphological preservation that have limited traditional approaches. Through its proprietary double-Z probe design and branched amplification system, RNAscope delivers single-molecule sensitivity while maintaining exceptional specificity and tissue morphology. Quantitative comparisons demonstrate strong correlation with qRT-PCR, and specialized applications like intronic probing enable previously challenging investigations in fields like cardiac regeneration. As spatial transcriptomics continues to evolve, RNAscope's robust methodology provides researchers with a reliable tool for precise gene expression analysis within native tissue contexts, effectively balancing the critical demands of detection sensitivity and morphological preservation that underlie rigorous spatial genomics research.
Evidence-Based Comparison: How RNAscope Stacks Up Against Gold Standard Techniques
The accurate detection and localization of RNA biomarkers are fundamental to advancing our understanding of disease mechanisms and developing targeted therapies. For decades, quantitative PCR (qPCR) and quantitative reverse-transcription PCR (qRT-PCR) have served as cornerstone techniques for gene expression analysis due to their sensitivity and quantitative capabilities [16]. Similarly, immunohistochemistry (IHC) has remained the gold standard for protein visualization in tissue context. However, these methods present significant limitations—PCR-based techniques lack spatial context as they require tissue homogenization, while IHC depends on antibody availability and quality and does not directly measure RNA expression [16] [49]. RNAscope, a novel RNA in situ hybridization (ISH) platform, addresses these limitations by enabling single-molecule RNA visualization within intact tissue morphology [2]. This systematic review synthesizes evidence on the concordance between RNAscope and traditional methods, positioning it as a powerful complementary technique in the researcher's arsenal for precise biomarker detection and validation.
RNAscope Technology: Principles and Workflow
Core Technological Principle
RNAscope employs a proprietary probe design strategy that differentiates it from traditional RNA ISH methods. The technology utilizes paired "Z" probes that specifically hybridize to the target RNA sequence [16]. Each probe set consists of three elements: a target-hybridizing region, a spacer sequence, and a tail that binds to pre-amplifier molecules [16]. This unique design requires two independent probes to bind adjacent sites on the target RNA before signal amplification can proceed, ensuring exceptional specificity by minimizing non-specific binding and background noise [2]. The mechanism allows for single-RNA-molecule detection at single-cell resolution while preserving tissue morphology, bridging a critical gap between molecular detection and histological context [54].
Signal Amplification and Visualization
The signal amplification system of RNAscope provides significant sensitivity advantages over traditional approaches. After the "Z" probes hybridize to the target RNA, a multi-step amplification cascade begins: (1) pre-amplifier molecules attach to the probe tails, (2) multiple amplifier sequences bind to each pre-amplifier, and (3) enzyme-conjugated label probes bind to the amplifiers, enabling visual detection [16]. This sophisticated process results in up to 8,000-fold signal amplification, allowing routine detection of individual RNA molecules despite potential RNA degradation in formalin-fixed, paraffin-embedded (FFPE) tissues [16] [2]. Detection can be achieved through either chromogenic substrates for bright-field microscopy or fluorescent labels for multiplex analysis, providing flexibility for various research applications and imaging platforms.
Standardized Workflow and Quality Control
The RNAscope workflow begins with slide preparation from FFPE tissues, tissue microarrays, fresh frozen tissues, or fixed cells [16]. Prepared slides then undergo three key procedural steps: permeabilization, hybridization, and signal amplification. Notably, these steps can be performed manually or automated for high-throughput applications [16]. A critical strength of the RNAscope system is its integrated quality control, employing negative control probes targeting the bacterial dapB gene to confirm absence of background noise, and positive control probes for housekeeping genes (PPIB, Polr2A, or UBC) to verify RNA integrity and assay performance [16]. This rigorous quality framework ensures reliable, interpretable results across different tissue types and experimental conditions.
Table 1: Essential Research Reagent Solutions for RNAscope Implementation
| Reagent/Category | Specific Examples | Function and Application |
|---|---|---|
| Probe Types | Target-specific "Z" probes, Positive control probes (PPIB, Polr2A, UBC), Negative control probe (dapB) | Target RNA detection; assay validation and quality control; background assessment |
| Sample Processing | FFPE tissue sections, Fresh frozen tissues, Fixed cells | Tissue preservation and preparation for RNAscope analysis |
| Detection Systems | Chromogenic dyes, Fluorescent dyes | Signal visualization for bright-field or fluorescence microscopy |
| Analysis Platforms | Halo, QuPath, Aperio software | Automated quantification of RNA expression and localization |
Diagram 1: RNAscope workflow and quality control framework. The process integrates standardized procedural steps with rigorous quality assessment to ensure reliable RNA detection.
Concordance with Gold Standard Methods: Systematic Evidence
RNAscope vs. PCR-based Methods
A systematic review conducted in 2021 evaluated RNAscope performance compared to established molecular techniques, including qPCR and qRT-PCR [16]. The analysis encompassed 27 retrospective studies, primarily focusing on cancer samples, and demonstrated consistently high concordance rates between RNAscope and PCR-based methods, ranging from 81.8% to 100% [16]. This strong agreement confirms that RNAscope maintains the detection accuracy of gold standard molecular techniques while adding the crucial dimension of spatial context. The high sensitivity and specificity of RNAscope, both potentially reaching 100%, stem from its unique signal amplification system and stringent probe design requirements that minimize background noise and false-positive signals [16] [2].
The exceptional concordance is particularly notable given the fundamental methodological differences between these techniques. While qPCR and qRT-PCR require RNA extraction from homogenized tissues, averaging expression across cell populations, RNAscope preserves tissue architecture and enables single-cell resolution analysis [16]. This distinction makes RNAscope particularly valuable for detecting heterogeneous gene expression within complex tissues and identifying rare cell populations that might be diluted beyond detection in bulk PCR analyses. The technique's ability to reliably work with partially degraded RNA from FFPE specimens further enhances its utility for clinical research and biomarker validation on archival samples [16].
RNAscope vs. Immunohistochemistry
The same systematic review reported that RNAscope's concordance with IHC was somewhat more variable, ranging from 58.7% to 95.3% [16]. This wider range reflects the fundamental difference in what each technique measures—RNAscope detects RNA transcripts while IHC detects protein expression. The discordance can be attributed to several biological and technical factors, including post-transcriptional regulation, differences in protein and RNA turnover rates, and variability in IHC antibody performance [16] [14]. Importantly, RNAscope does not aim to replace IHC but rather serves as a complementary approach that can provide additional layers of molecular information.
In many cases, RNAscope offers advantages over IHC for specific applications. For viral detection, RNAscope provides greater sensitivity and specificity compared to IHC, and does not require custom antibody development [49]. For biomarkers with limited antibody availability or quality, RNAscope enables direct RNA visualization that can confirm IHC findings or resolve equivocal results. The combination of both techniques on the same tissue section provides a comprehensive view of gene expression from transcription to translation, offering powerful insights into disease mechanisms and treatment responses [16].
Table 2: Concordance Rates Between RNAscope and Established Methods
| Comparison Method | Concordance Range | Key Factors Influencing Concordance | Advantages of RNAscope |
|---|---|---|---|
| qPCR/qRT-PCR | 81.8% - 100% [16] | RNA preservation, tumor cellularity | Spatial context preservation, single-cell resolution, works with FFPE tissues |
| DNA In Situ Hybridization | High concordance (specific rates not provided) [16] | Probe specificity, amplification efficiency | Single-molecule sensitivity, higher throughput capability |
| Immunohistochemistry | 58.7% - 95.3% [16] | Post-transcriptional regulation, antibody quality | Direct RNA detection, no antibody requirements, superior sensitivity for some targets |
Experimental Protocols for Method Comparison
Sample Preparation and Nucleic Acid Extraction
Robust comparison studies employ standardized sample processing protocols to ensure valid method comparisons. For FFPE tissues, researchers typically cut sequential sections (5μm for DNA/RNA extraction, 5μm for IHC, and 4-5μm for FISH) to enable parallel analysis of the same tumor regions [78]. Before molecular analysis, hematoxylin and eosin-stained sections are examined by a pathologist to determine tumor cellularity, with samples below 70% tumor content often undergoing macro-dissection or laser capture microdissection to enrich malignant cells [78]. DNA and RNA are commonly extracted using commercial kits (e.g., QIAamp DNA FFPE Tissue Kit, Paradise Reagent System) with inclusion of DNase I treatment for RNA preparations [78]. Nucleic acid quantity and quality are assessed using both spectrophotometric (NanoDrop) and fluorimetric methods (Qubit), with the latter providing more accurate quantification for downstream molecular applications [78].
RNAscope Procedure and Detection
The RNAscope protocol follows a standardized three-step process after slide preparation [16]. First, tissues undergo controlled permeabilization to enable probe access while preserving RNA integrity. Next, target-specific "Z" probes are hybridized to the RNA of interest. Finally, the signal amplification cascade is performed according to manufacturer specifications. For multiplex analysis, sequential hybridization and amplification steps can be performed with different probe sets [16]. Detection utilizes either chromogenic substrates for bright-field microscopy or fluorescent labels for fluorescence imaging. Appropriate controls must be included in each run: positive control probes for housekeeping genes to confirm RNA integrity and assay performance, and negative control probes (bacterial dapB gene) to verify absence of background signal [16].
Analysis and Quantification Methods
RNAscope results are quantified by counting the number of distinct dots per cell, with each dot representing an individual RNA molecule [16]. Analysis can be performed manually by trained personnel or using automated image analysis software such as Halo, QuPath, or Aperio [16] [52]. These platforms enable quantitative assessment of RNA expression levels while documenting spatial distribution patterns within tissue architecture. For PCR-based comparisons, qRT-PCR is typically performed using validated primer sets and reference genes (e.g., ACTB, RPLP0, TFRC) on compatible real-time PCR systems [78] [79]. Normalization strategies include the delta-delta CT method or quantile normalization when comparing across different sample sets [79].
Diagram 2: Method comparison highlighting what each technique detects and their concordance relationships. RNAscope bridges the gap between spatial context and molecular detection.
Discussion and Clinical Implications
Complementary Role in Diagnostic Resolution
The evidence synthesized in this review supports RNAscope as a valuable complementary technique rather than a wholesale replacement for existing methods. The high concordance with PCR-based techniques validates its quantitative accuracy, while the spatial information it provides addresses a critical limitation of bulk RNA analysis [16]. For IHC, RNAscope serves as an orthogonal method that can resolve equivocal cases, particularly when antibody performance is suboptimal or when investigating genes with known post-transcriptional regulation [16] [14]. This complementary relationship enhances diagnostic confidence and enables more precise patient stratification in both research and clinical settings.
RNAscope demonstrates particular utility in specific challenging scenarios: identifying heterogeneous gene expression within tumors, detecting low-abundance transcripts, validating RNA biomarkers in FFPE tissues, and detecting viral RNAs in infectious disease research [16] [49]. Its ability to work reliably with FFPE tissues makes it exceptionally valuable for retrospective studies utilizing archival clinical samples with associated outcome data [2]. The technology's capacity for multiplex analysis further enables investigation of gene co-expression patterns and cell-cell interactions within the tissue microenvironment, providing insights that would require multiple parallel experiments with traditional methods [16].
Technical Considerations and Implementation Challenges
Despite its advantages, RNAscope implementation presents certain practical considerations. The technique requires specialized equipment for automated processing and high-quality imaging systems for proper quantification [16]. Analysis can be time-consuming without automated image analysis solutions, and optimal results depend on proper tissue fixation and processing [16]. The requirement for RNA preservation in tissue samples necessitates careful attention to pre-analytical variables, though RNAscope has demonstrated robustness with partially degraded RNA from archival FFPE blocks [2]. Additionally, while RNAscope probes are available for an expanding list of targets, custom probe development may be necessary for novel targets, though the turnaround time of approximately two weeks is relatively rapid [49].
From a cost perspective, RNAscope may represent a more significant initial investment than traditional PCR or IHC, but this must be balanced against the rich spatial information obtained and the potential to reduce confirmatory testing through higher specificity. For clinical applications, further validation studies following regulatory guidelines would be necessary to establish standardized scoring thresholds and demonstrate clinical utility for specific indications [16]. As with any emerging technology, establishment in routine diagnostics will require technical standardization, training programs, and consensus guidelines for interpretation and reporting.
RNAscope technology represents a significant advancement in molecular pathology, offering unprecedented ability to visualize RNA expression within morphological context. The substantial concordance with qPCR and qRT-PCR (81.8-100%) validates its analytical performance, while its complementary relationship with IHC enables more comprehensive biomarker assessment [16]. As research increasingly focuses on spatial biology and heterogeneous tissue environments, RNAscope provides a crucial methodological bridge between bulk molecular analysis and tissue morphology. Its robust performance with FFPE tissues positions it as particularly valuable for translational research utilizing clinical archives. While further standardization and validation will enhance its clinical adoption, RNAscope already offers researchers a powerful tool to address fundamental questions in gene expression regulation, disease pathogenesis, and treatment response mechanisms across diverse fields from oncology to infectious disease.
In modern molecular pathology and drug development, accurately measuring biomarker expression is fundamental for both diagnostics and therapeutic decision-making. The central challenge researchers face is the frequent discrepancy between detecting a biomarker's RNA transcripts and its translated protein. Immunohistochemistry (IHC), which detects proteins using antibodies, remains the gold standard for many clinical applications due to its ability to visualize protein localization within tissue architecture. However, emerging RNA in situ hybridization technologies, particularly RNAscope, offer unprecedented sensitivity for RNA detection while maintaining morphological context. This comparison guide objectively evaluates these complementary technologies, examining the root causes of discordant results and providing researchers with methodological frameworks to resolve these critical discrepancies.
The fundamental relationship between RNA transcription and protein translation is complex and regulated by numerous factors including post-transcriptional modifications, translation efficiency, and protein degradation rates. While mRNA levels often correlate with protein abundance, this relationship is imperfect, leading to challenging interpretive scenarios in both research and clinical settings. Understanding the technical and biological factors underlying these discrepancies is essential for advancing precision medicine and developing more accurate diagnostic paradigms.
Technical Principles: Fundamental Differences in Detection
Immunohistochemistry (IHC)
IHC operates on the principle of antibody-antigen recognition, utilizing labeled antibodies to detect specific protein epitopes within tissue sections [80]. The process begins with antigen retrieval to unmask epitopes altered by tissue fixation, followed by sequential application of primary antibodies, secondary detection systems, and chromogenic or fluorescent visualization [80]. This methodology provides direct evidence of protein presence, localization, and relative abundance, making it invaluable for assessing functional gene products. However, IHC suffers from limitations including antibody specificity issues, batch-to-batch variability, and subjective scoring systems that contribute to interobserver variability [81] [82].
RNAscope In Situ Hybridization
RNAscope represents a significant advancement over traditional in situ hybridization methods, employing a novel double-Z probe design that enables single-molecule RNA visualization while preserving tissue morphology [16] [1]. The proprietary probe design features two separate "Z probes" that must bind adjacent to each other on the target RNA sequence to initiate a signal amplification cascade [16]. This requirement for dual recognition dramatically reduces non-specific binding and background noise, addressing the primary limitations of conventional ISH methods. The amplification system enables visualization of individual RNA molecules as distinct dots, allowing for both qualitative localization and quantitative assessment [16].
Table 1: Core Technological Principles Comparison
| Feature | Immunohistochemistry (IHC) | RNAscope ISH |
|---|---|---|
| Detection Target | Protein epitopes | RNA molecules |
| Signal Mechanism | Antibody-antigen binding | Nucleic acid hybridization |
| Key Reagents | Primary and secondary antibodies | ZZ probe pairs, amplification system |
| Visualization | Chromogenic or fluorescent deposition | Chromogenic or fluorescent dots |
| Cellular Resolution | Protein localization within subcellular compartments | RNA localization within cytoplasm/nucleus |
| Specificity Control | Isotype controls, absorption tests | Negative control probes (e.g., bacterial dapB) |
Diagram 1: Comparative Workflows of IHC and RNAscope Technologies
Quantitative Performance Comparison
Analytical Sensitivity and Specificity
Direct head-to-head comparisons reveal significant differences in the performance characteristics of IHC versus RNAscope. In a comprehensive study detecting SARS-CoV-2 in human tissues, RNAscope demonstrated 86.7% sensitivity and 100% specificity when compared with quantitative RT-PCR as the gold standard [83]. In contrast, IHC targeting the SARS-CoV nucleocapsid protein showed similar sensitivity (85.7%) but markedly reduced specificity (53.3%), with false positive staining observed in 5 of 13 control cases [83]. This specificity advantage of RNAscope was further reflected in interobserver agreement, which ranged from "moderate to almost perfect" for ISH compared to only "slight to moderate" for IHC among four independent pathologists [83].
The high specificity of RNAscope stems from its unique probe design requirement for dual probe binding, which dramatically reduces off-target hybridization [16]. This technological advantage becomes particularly important when detecting targets with closely related family members or when working with partially degraded samples, where antibody cross-reactivity can lead to misleading IHC results.
Concordance Rates Across Biomarker Types
A systematic review comparing RNAscope with gold standard techniques across 27 studies reported high concordance rates between RNAscope and PCR-based methods (81.8-100%) but more variable concordance with IHC (58.7-95.3%) [16]. This discrepancy highlights the fundamental difference between detecting RNA transcripts versus protein products, which can be influenced by post-transcriptional regulation, translation efficiency, and protein turnover rates.
In cancer biomarker applications, a recent study investigating HER2-low breast cancer found that while mean rank HER2 mRNA levels were significantly higher in HER2-low cases compared to HER2 0/ultralow groups, substantial discordance existed at the individual case level [81]. Specifically, 58.8% of cases classified as HER2 0/ultralow by IHC were classified as HER2-low by MammaTyper RT-qPCR, while 2.8% showed the opposite pattern [81]. These findings underscore the complex relationship between mRNA abundance and protein expression for therapeutic targets.
Table 2: Quantitative Performance Metrics Across Studies
| Study/Application | Sensitivity | Specificity | Concordance with Gold Standard | Interobserver Agreement |
|---|---|---|---|---|
| SARS-CoV-2 Detection [83] | 86.7% (ISH)85.7% (IHC) | 100% (ISH)53.3% (IHC) | qRT-PCR reference | Moderate-almost perfect (ISH)Slight-moderate (IHC) |
| Systematic Review [16] | N/A | N/A | 81.8-100% (vs. PCR)58.7-95.3% (vs. IHC) | Not reported |
| HER2-low BC [81] | N/A | N/A | 58.8% discordance rate(IHC 0 vs. mRNA-low) | Limited IHC agreement |
Methodological Considerations
Experimental Protocols for Comparative Studies
For researchers designing head-to-head comparison studies, specific methodological details critically impact result interpretation. In the SARS-CoV-2 study, both IHC and RNAscope were performed on consecutive formalin-fixed paraffin-embedded (FFPE) tissue sections from COVID-19 autopsies [83]. The IHC protocol utilized a primary antibody against SARS-CoV nucleocapsid protein with heat-induced epitope retrieval, while RNAscope employed target probes against SARS-CoV-2 RNA with protease digestion for tissue permeabilization [83].
For biomarker studies, the recent investigation comparing RNA sequencing with IHC employed rigorous validation protocols [14]. IHC staining was performed using an automated stainer with specific primary antibodies according to manufacturing guidelines, validated as laboratory-developed tests [14]. Slides were scanned at 20× magnification and quantified using QuPath software with positive cell detection algorithms for nuclear markers, while membrane markers like HER2 and PD-L1 were scored visually by two pathologists according to clinical guidelines [14].
Technical Advantages and Limitations
Each technology presents distinct advantages that make it suitable for specific applications. RNAscope offers superior specificity and the ability to detect virtually any RNA sequence with appropriate probe design, including targets for which high-quality antibodies are unavailable [82]. The capacity for multiplexing to detect several RNA targets simultaneously and precise single-cell resolution without signal diffusion represents additional advantages for complex tissue analysis [16].
IHC maintains the advantage of directly detecting functional gene products (proteins) and established clinical utility for numerous biomarkers [80]. The widespread availability of automated platforms, extensive validation history, and familiarity among pathologists make IHC the more practical choice for routine clinical applications in many settings. However, challenges with antibody validation, batch-to-batch variability, and subjective interpretation remain significant limitations [82].
Diagram 2: Comparative Advantages and Limitations of IHC and RNAscope
Research Reagent Solutions
Successful implementation of either technology requires appropriate selection of reagents and controls. The following table outlines essential research reagents for both platforms:
Table 3: Essential Research Reagents and Controls
| Reagent Category | Specific Examples | Function and Importance |
|---|---|---|
| IHC Primary Antibodies | Anti-SARS-CoV nucleocapsid [83], Anti-HER2 [81], Anti-PD-L1 [14] | Target-specific binding; clone selection critical for specificity |
| IHC Detection Systems | Polymer-based HRP systems [80], SAVIEW PLUS HRP [80] | Signal amplification and visualization; reduces background |
| RNAscope Probes | SARS-CoV-2 target probes [83], PPIB, Polr2A, UBC [16] | Target RNA hybridization; control probes validate assay performance |
| RNAscope Amplification | PreAmplifier, Amplifier [16], Label Probes [1] | Signal amplification cascade enabling single-molecule detection |
| Control Reagents | Positive control tissues [84], dapB negative control [16], Isotype controls | Assay validation and specificity verification |
Resolution Strategies for Discordant Results
Biological Versus Technical Discrepancies
When facing discordant RNA and protein detection results, researchers must systematically evaluate whether discrepancies reflect biological reality or technical artifacts. Biological discrepancies occur when genuine dissociations exist between mRNA transcription and protein translation due to:
- Post-transcriptional regulation (microRNAs, RNA binding proteins)
- Altered translation efficiency
- Impaired protein folding or degradation
- Differential protein trafficking
Technical discrepancies arise from methodological limitations:
- Suboptimal antibody specificity in IHC [82]
- RNA degradation affecting ISH sensitivity [16]
- Improper epitope retrieval or fixation artifacts
- Threshold setting for positivity determination
Integrated Validation Framework
A sequential validation approach resolves discordant results effectively. First, verify technical quality using appropriate controls: RNA integrity for RNAscope (housekeeping genes like PPIB) [16] and antibody specificity for IHC (isotype controls, absorption tests) [84]. Second, employ orthogonal validation methods such as qRT-PCR to confirm RNAscope results [83] or Western blotting to verify IHC findings. Third, consider biological context including tissue type, cellular heterogeneity, and expected subcellular localization patterns.
For targets with persistent discrepancies despite technical validation, the most biologically plausible explanation should guide interpretation. In the case of HER2-low breast cancer, the substantial proportion of cases classified as HER2 0/ultralow by IHC but HER2-low by mRNA expression may represent either technical IHC limitations or genuine biological states with post-transcriptional regulation [81]. Clinical outcome correlation ultimately determines which detection method better predicts therapeutic response.
The head-to-head comparison between IHC and RNAscope reveals complementary strengths rather than absolute superiority of either platform. IHC remains indispensable for direct protein detection with established clinical utility, while RNAscope offers superior specificity, single-molecule sensitivity, and the ability to detect targets without available antibodies. The observed discrepancies between RNA and protein detection frequently reflect both technical limitations and genuine biological phenomena rather than mere methodological failure.
Researchers and drug development professionals should select detection methods based on specific application requirements, recognizing that orthogonal verification using both platforms provides the most comprehensive biomarker assessment. As novel therapeutic modalities increasingly target specific RNA species or require precise quantification of low-abundance targets, RNAscope offers a powerful complementary tool to traditional IHC. Future diagnostic paradigms will likely integrate both technologies to leverage their respective advantages, ultimately enhancing precision medicine across oncology, infectious disease, and broader pathology practice.
In situ hybridization (ISH), a foundational technique for visualizing nucleic acids within their native cellular and tissue contexts, has undergone revolutionary advancements to overcome the limitations of conventional methods, particularly for detecting low-abundance transcripts. While traditional ISH methods, often relying on digoxigenin (DIG)-labeled RNA probes, can achieve good sensitivity with optimized protocols, they frequently struggle with complexity, time-consuming procedures, and difficulty in multiplexing [85]. The emergence of highly sensitive ISH variants, including the commercialized RNAscope and open-design methods like Hybridization Chain Reaction (HCR), clampFISH, and SABER FISH, has redefined the limits of RNA detection, enabling single-molecule resolution and robust multiplexing [85]. This guide provides an objective, data-driven comparison of these advanced ISH platforms, framing their performance within the broader thesis of how RNAscope's standardized detection system contrasts with the customizable, researcher-driven approaches of other sensitive variants. Understanding the principles, performance metrics, and practical considerations of each method is crucial for researchers and drug development professionals to select the optimal tool for their specific biological questions and resource constraints.
Principles and Signal Amplification Mechanisms
The high sensitivity of modern ISH methods stems from sophisticated signal amplification strategies that move beyond simple enzymatic detection. These methods generally share a two-step procedure: (1) the hybridization of synthetic, target-specific primary probes, and (2) the binding of multiple secondary molecules to these probes, resulting in a massive accumulation of signal at the site of each target RNA molecule [85].
The table below summarizes the core amplification principles and key structural features of each method.
Table 1: Core Principles and Probe Design of Sensitive ISH Variants
| Method | Signal Amplification Principle | Probe Structure | Key Design Feature |
|---|---|---|---|
| RNAscope | Proprietary branched DNA (bDNA) assay with sequential hybridization of pre-amplifier and amplifier molecules [85] [2]. | Multiple pairs of "ZZ" probes binding adjacent sites on the target [2] [54]. | Patented "double Z" probe design for background suppression and single-molecule visualization [2] [54]. |
| HCR (Hybridization Chain Reaction) | Enzyme-free, isothermal amplification using metastable DNA hairpins that polymerize via a chain reaction initiated by the primary probe [85] [86]. | Paired probes that, when bound adjacently, expose an initiator sequence [85] [86]. | Nonlinear hybridization chain reaction; amplification time can be adjusted by user [85]. |
| clampFISH | Click chemistry-assisted ligation of padlock probes to form circular structures, followed by multiple hybridizations of fluorescent reporters to the locked probe [85]. | Padlock probes that hybridize to form a circular structure [85]. | Probes are fixed to the target via ligation, enhancing specificity and signal stability [85]. |
| SABER FISH (Signal Amplification By Exchange Reaction) | Primer exchange reaction (PER) to enzymatically synthesize long concatemers on the primary probe, providing many binding sites for fluorescently labeled imager strands [85] [87]. | Primary probes extended with a uniform repeating sequence (concatemer) of customizable length [85] [87]. | Amplification degree is tunable by varying concatemer length during synthesis [85]. |
To illustrate the logical workflow and decision points involved in selecting and applying these techniques, the following diagram maps the path from experimental goal to method selection and outcome.
Direct Performance Comparison: Quantitative and Qualitative Metrics
A critical step in benchmarking is the direct comparison of technical performance. Recent studies have provided quantitative data on sensitivity, signal strength, and processing speed, allowing for an objective evaluation.
Key Performance Metrics Table
Table 2: Experimental Performance Benchmarking of Sensitive ISH Methods
| Method | Detection Sensitivity | Signal Strength & Resolution | Processing Time (per round) | Minimum Probe Pairs |
|---|---|---|---|---|
| RNAscope | Single RNA molecule detection [2] [54]. | High signal-to-noise; granular signals for single-molecule resolution [85] [2]. | ~1 day for full assay [85]. | Not specified; proprietary design. |
| HCR FISH | Single RNA molecule detection [85]. | Strong signal; user-defined amplification time [85] [86]. | ≥8 hours for amplification [88] [86]. | 3 pairs for robust signal [86]. |
| SABER FISH | Single RNA molecule detection [85]. | Strong, tunable signal via concatemer length [85] [87]. | 1-3 days for full assay [85]. | Varies with target and concatemer length. |
| clampFISH | Single RNA molecule detection [85]. | High specificity due to ligation; granular signals [85]. | 1-3 days for full assay [85]. | Not explicitly stated. |
| TDDN-FISH | Single RNA molecule detection, including short RNAs (e.g., miRNA) [88]. | Significantly stronger than smFISH and HCR FISH [88]. | ~1 hour post-hybridization [88]. | 3 probes for strong signal on mRNA [88]. |
Emerging Methods and Enhanced Protocols
The field continues to evolve with new methods offering further improvements. TDDN-FISH (Tetrahedral DNA Dendritic Nanostructure–Enhanced FISH), for instance, uses self-assembling 3D DNA nanostructures for signal amplification. This enzyme-free method is reported to be approximately eightfold faster per round than HCR-FISH and generates stronger signals than single-molecule FISH (smFISH), enabling the detection of short RNAs [88].
Furthermore, established methods are being refined into more versatile platforms. The OneSABER framework demonstrates how SABER-derived probes can be unified with various detection modalities, including canonical colorimetric assays, Tyramide Signal Amplification (TSA), and HCR, creating a "one probe fits all" approach that increases flexibility and reduces costs [87].
Similarly, the Yn-situ method incorporates a Y-branched preamplifier probe that binds to target probes and triggers multiple HCR reactions. This design enhances sensitivity, allowing for robust detection with as few as three probe pairs compared to the 20-probe sets often required in standard HCR, thereby reducing cost and complexity [86].
Experimental Design and Protocol Considerations
The practical implementation of these methods varies significantly, impacting the required expertise, time, and cost. A detailed understanding of the experimental workflow is essential for successful adoption.
Comparative Workflow and Protocol Table
Table 3: Experimental Workflow and Resource Requirements Comparison
| Method | Probe Design & Synthesis | Key Steps in Protocol | Combination with Immunostaining | Multiplexing Capability |
|---|---|---|---|---|
| RNAscope | Provided by manufacturer; proprietary [85] [49]. | Standardized, simplified workflow with pre-optimized reagents; often a single-day protocol [85] [2]. | Excellent; low hybridization temperatures preserve antigens [85]. | Easy; commercial kits available for duplex and multiplex assays [49] [54]. |
| HCR FISH | Designed by user; can be outsourced for synthesis [85] [86]. | Hybridization of target probes, followed by amplification with DNA hairpins for a defined time [85] [86]. | Excellent; enzyme-free and low-temperature hybridization [85]. | Easy; sequential or parallel HCR with orthogonal hairpins is feasible [85] [87]. |
| SABER FISH | Designed by user; concatemers synthesized in vitro via Primer Exchange Reaction (PER) [85] [87]. | Probe extension (PER), hybridization, then binding of fluorescent imager strands [85] [87]. | Excellent [85]. | Easy; supported by combinatorial barcoding [87]. |
| clampFISH | Designed by user [85]. | Hybridization of padlock probes, ligation, then multiple rounds of reporter hybridization [85]. | Excellent [85]. | Easy [85]. |
The following diagram visualizes the core signal amplification pathways for RNAscope, HCR, and SABER FISH, highlighting their distinct mechanistic approaches.
Key Research Reagent Solutions
The successful execution of these advanced ISH protocols relies on a set of crucial reagents and materials. The following table details these essential components and their functions.
Table 4: Essential Reagents and Materials for Sensitive ISH Methods
| Reagent / Material | Function | Method Applicability |
|---|---|---|
| Primary Probes | Synthetic oligonucleotides that bind specifically to the target RNA sequence. | All methods (RNAscope, HCR, SABER, clampFISH) [85]. |
| Signal Amplifiers | Branched DNA structures (RNAscope), DNA hairpins (HCR), or concatemers (SABER) that bind to primary probes to amplify signal. | Specific to each method's principle [85] [86] [87]. |
| Fluorophore-Labeled Reporters | Oligonucleotides or tyramides conjugated to fluorescent dyes for signal detection. | All fluorescent methods [85] [87]. |
| Formamide-Based Hybridization Buffer | Creates denaturing conditions that facilitate specific probe binding to the target RNA. | All methods [85] [86]. |
| Protease or Permeabilization Reagents | (e.g., Proteinase K) Treat tissues to allow probe penetration; note RNAscope's newer protease-free options [89]. | Most methods, though requirements vary [85] [89]. |
| EDC (1-ethyl-3-(3-dimethylaminopropyl) carbodiimide) | A carbodiimide fixative that crosslinks RNA to proteins, reducing degradation and improving signal quality, especially in stored tissues. | Can be applied as a pre-treatment to enhance any method's robustness [86]. |
Discussion: Strategic Selection of ISH Methods
The choice between RNAscope and other sensitive ISH variants is not a matter of absolute superiority but rather strategic alignment with research goals, resources, and sample types.
Integrated Cost-Benefit Analysis
A critical factor in method selection is a holistic consideration of costs, which encompasses not only monetary expenses but also time investment and expertise.
Table 5: Comprehensive Cost-Benefit Analysis of Sensitive ISH Platforms
| Method | Monetary Cost | Time & Expertise Cost | Ideal Use Case Scenarios |
|---|---|---|---|
| RNAscope | High cost per sample; cost increases proportionally with sample number [85]. | Low time cost; minimal optimization needed; easy to learn [85]. | - Clinical diagnostics and pathology [2] [90].- Projects with limited scope and need for rapid results.- Labs lacking resources for extensive protocol development. |
| HCR, SABER, clampFISH | Moderate initial setup; cost per sample decreases with increasing number of samples [85]. | High time cost; requires significant optimization and expertise in probe design [85]. | - Large-scale screening studies.- Highly multiplexed experiments requiring customization [87].- Detection of short transcripts or novel targets with user-defined parameters.- Academic labs prioritizing lower long-term costs and open platforms. |
The benchmarking data clearly illustrates a trade-off between standardization and customization. RNAscope stands out for its operational simplicity, reliability, and rapid turnaround, making it an powerful tool for diagnostic applications and targeted studies where cost is a secondary concern. Its recent integration into automated platforms and development of protease-free assays further solidifies its position in clinical and translational research landscapes [89].
In contrast, HCR FISH, SABER FISH, and clampFISH offer a different value proposition: flexibility and control. These open-design platforms empower researchers to tune sensitivity, customize multiplexing panels, and ultimately reduce per-sample costs for large projects, albeit at the expense of initial development time and expertise. The emergence of unified platforms like OneSABER [87] and enhanced methods like Yn-situ [86] and TDDN-FISH [88] demonstrates a vigorous trend towards more accessible, efficient, and powerful open-approach ISH technologies.
For the researcher, the decision matrix is clear. Prioritize time-to-result and ease-of-use for focused studies: choose RNAscope. Prioritize long-term cost-efficiency, high multiplexing, and customizability for large-scale discovery projects: invest in the development of HCR, SABER, or related open-platform methods.
The integration of precision medicine into clinical practice hinges on reliable biomarkers to guide therapeutic decisions. In situ RNA analysis is highly desirable in molecular pathology as it allows for the examination of biomarker status within the full histopathological context of clinical specimens. However, its clinical adoption has been limited by the technical complexity, insufficient sensitivity, and high background of traditional RNA in situ hybridization (ISH) techniques. RNAscope technology, a novel RNA ISH platform, addresses these limitations through a unique probe design that enables single-molecule visualization while preserving tissue morphology. This case study details the CLIA-validation of a specific RNAscope assay for Dickkopf-1 (DKK1) in gastric and gastroesophageal junction (G/GEJ) adenocarcinoma, providing a template for the development of robust in situ RNA-based clinical assays [50] [2].
RNAscope Technology: Principles and Advantages Over Traditional Methods
Core Technology and Mechanism
RNAscope is based on a patented signal amplification and background suppression technology. Unlike traditional RNA ISH, which uses a single probe conjugated to a label, RNAscope employs a pair of so-called "Z" probes that hybridize to the target RNA [16]. This design is the cornerstone of its performance:
- High Specificity: The requirement for two independent "Z" probes to bind in close proximity for signal amplification to occur suppresses off-target binding and background noise. The specificity can reach 100% [16].
- High Sensitivity: A multi-step amplification process follows probe binding. Each target RNA molecule can bind multiple "Z" probe pairs, each of which can subsequently bind pre-amplifiers and amplifiers, culminating in the attachment of up to 400 labeled probes per "Z" dimer. This allows for single RNA molecule detection with a signal amplification of up to 8,000 times [16].
The following diagram illustrates the key principle of the RNAscope assay:
Performance Comparison with Traditional Techniques
A 2021 systematic review compared RNAscope to established "gold standard" methods for gene expression analysis. The results, summarized in the table below, confirm its superior performance characteristics [16].
Table 1: RNAscope Performance Compared to Gold Standard Techniques
| Technique | Concordance Rate with RNAscope | Key Advantages of RNAscope | Key Limitations of Comparison Method |
|---|---|---|---|
| IHC | 58.7% - 95.3% | Measures RNA directly; higher sensitivity for low-abundance targets; avoids antibody cross-reactivity. | Measures protein; discordance often due to post-transcriptional regulation. |
| qPCR / qRT-PCR | 81.8% - 100% | Preserves tissue morphology and spatial context; allows cellular heterogeneity assessment. | Requires RNA extraction ("grind-and-bind"); loses histological context. |
| DNA ISH | 81.8% - 100% | Detects expressed RNA, not genomic DNA. | Detects gene presence/amplification, not expression. |
This data demonstrates that RNAscope is a reliable and robust method that complements and, in some cases, surpasses traditional techniques, particularly IHC. The review concluded that RNAscope could be used to confirm unclear results from standard methods [16]. In the context of biomarker development for targeted therapies, RNAscope assays have been reported to be "more sensitive than IHC and often correlate better with response to treatment," as less sensitive IHC assays can report false-negative results [91].
CLIA-Validation of a DKK1 RNAscope Assay: A Detailed Case Study
Clinical Background and Rationale
Dickkopf-1 (DKK1) is a secreted modulator of Wnt signaling that is frequently overexpressed in tumors and is associated with poor clinical outcomes [50]. DKN-01 is a humanized monoclonal therapeutic antibody that binds DKK1 with high affinity. A clinical hypothesis was formed that tumoral DKK1 expression could serve as a predictive biomarker for patient response to DKN-01 [50]. Initial retrospective analysis of tumor tissues from a phase 1b/2a study showed that G/GEJ cancer patients with elevated tumoral DKK1 mRNA levels were more likely to benefit from a combination of DKN-01 and pembrolizumab [50]. This finding necessitated the development and validation of a robust assay to prospectively identify patients with high DKK1 expression for subsequent clinical trials.
Assay Development and Initial Specificity/Sensitivity Assessment
The development of the DKK1 RNAscope chromogenic in situ hybridization (CISH) assay followed a structured pathway.
- Initial Assessment with Cell Lines: Researchers used four cancer cell lines (PC3, A549, HeLa, and Pfeiffer) with varying levels of DKK1 expression, as determined by RNA-Seq data from the Cancer Cell Line Encyclopedia (CCLE). The RNAscope results on a control formalin-fixed paraffin-embedded (FFPE) cell pellet array (CPA) showed a strong correlation with CCLE data, qPCR, and ELISA, confirming the assay's dynamic range and initial accuracy. The expected staining pattern was observed, with the strongest signal in PC3 cells and no signal in Pfeiffer cells [50].
- Specificity Testing: A key experiment involved testing a specificity CPA with cell lines expressing high levels of other Dickkopf family members (DKK2, DKK3, DKK4) but low DKK1. The DKK1 RNAscope probe showed minimal signal, demonstrating no cross-reactivity with related RNAs [50].
- Comparison with IHC: The RNAscope assay was compared to a DKK1 IHC assay. While both showed consistent results for high-expressing (PC3) and non-expressing (Pfeiffer) cell lines, RNAscope proved more sensitive, detecting DKK1 in HeLa cells where IHC signal was absent [50].
Validation According to CLIA Guidelines
The DKK1 RNAscope CISH assay was validated on 40 G/GEJ tumor resection specimens following CLIA guidelines. The validation assessed specificity, sensitivity, accuracy, and precision, and incorporated a digital image analysis algorithm (using QuPath software) to identify tumor cells and quantify the DKK1 signal, thereby reducing pathologist time and variability from manual scoring [50].
Table 2: Key Results from CLIA-Validation of the DKK1 RNAscope Assay
| Validation Parameter | Experimental Design & Samples | Key Outcome |
|---|---|---|
| Specificity | 40 G/GEJ tumor resections | DKK1 signal was localized specifically to tumor cells and was rarely detected in non-tumoral cells within the tissue. |
| Sensitivity | 40 G/GEJ tumor resections | The assay detected tumor cells with a wide range of DKK1 expression, including cells with a single dot (representing one RNA molecule). |
| Accuracy | Correlation with orthogonal methods | A significant correlation was observed between RNAscope digital H-scores and RNA-Seq data from 48 cell lines (Spearman’s rho = 0.86, p < 0.0001). |
| Precision | Inter-operator, inter-instrument, inter-day | The assay met all pre-defined acceptance criteria for precision (repeatability and reproducibility). |
| RNA Integrity | All samples with PPIB & dapB controls | All tumor resections showed adequate RNA integrity (PPIB signal ≥ 4 dots/cell) and acceptable background (minimal dapB signal). |
The validation was successful, and the assay, in conjunction with the digital image analysis solution, was deemed acceptable for the prospective screening of G/GEJ adenocarcinoma patients in a phase 2 clinical trial (NCT04363801) [50]. The following diagram illustrates this clinical validation workflow:
The Scientist's Toolkit: Essential Reagents and Controls for RNAscope
A properly controlled RNAscope experiment requires specific reagents and materials to ensure success.
Table 3: Essential Research Reagent Solutions for RNAscope Assays
| Item | Function & Importance | Examples / Notes |
|---|---|---|
| Target-Specific Probe Pairs | Designed to hybridize to the RNA of interest; the core of the assay's specificity. | Custom probes can be designed and manufactured in ~2 weeks [91]. |
| Positive Control Probes | Validate RNA integrity and assay performance. | Housekeeping genes: PPIB (moderate expression), POLR2A (low expression), UBC (high expression) [16] [6]. |
| Negative Control Probe | Assesses background noise and non-specific binding. | Bacterial gene dapB, which should not be present in human/animal samples [16] [6]. |
| HybEZ Oven / System | Maintains optimum humidity and temperature during hybridization steps; required for consistent results [6]. | |
| Specialized Slides & Mounting Media | Ensures tissue adhesion and signal preservation. | Superfrost Plus slides are required. Specific xylene-based or EcoMount/PERTEX mounting media must be used depending on the detection kit [6]. |
| Digital Image Analysis Software | Enables objective, quantitative scoring of dots per cell. | Halo, QuPath, Aperio [16] [50]. |
The successful CLIA-validation of the DKK1 RNAscope assay underscores its utility in the companion diagnostic (CDx) development pathway. This case study from Leap Therapeutics illustrates a common trajectory: starting with a Research Use Only (RUO) assay for hypothesis generation, progressing to a Laboratory Developed Test (LDT) for prospective patient screening in clinical trials, and ultimately planning for a transition to a fully approved in vitro diagnostic (IVD) [92]. This process requires close collaboration with regulatory agencies and partners, such as CLIA-certified laboratories, early in the drug development program [92].
In conclusion, the high sensitivity and specificity of RNAscope, combined with its ability to provide spatial context, make it a powerful tool for translating RNA biomarkers from research discoveries into clinically validated assays. The DKK1 case study provides a validated roadmap for this process, demonstrating that RNAscope technology can meet the rigorous standards of CLIA laboratories. This paves the way for its broader adoption in precision medicine, ensuring that the right patients are matched with the right targeted therapies based on the molecular profile of their tumors [50].
While numerous molecular techniques exist for nucleic acid and protein analysis, each possesses inherent strengths and limitations that dictate their appropriate application. RNAscope in situ hybridization has emerged as a powerful technology that occupies a unique niche within the molecular toolbox, complementing rather than replacing established methods. This review objectively examines how RNAscope's capacity for single-molecule RNA visualization within morphological context addresses specific gaps left by traditional in situ hybridization, immunohistochemistry, and PCR-based techniques. We present experimental data comparing performance metrics, detailed methodologies for implementation, and a structured framework for researchers to determine when RNAscope provides essential complementary data alongside established molecular approaches.
Molecular analysis in biomedical research relies on a diverse set of techniques, each optimized for specific types of information. Immunohistochemistry (IHC) provides protein localization within tissue architecture but depends on antibody availability and quality. PCR-based methods (qPCR, qRT-PCR) offer sensitive nucleic acid quantification but require tissue homogenization that eliminates spatial context. Traditional in situ hybridization (ISH) preserves spatial information but has historically suffered from limited sensitivity and specificity issues [16] [1]. The development of RNAscope represents a significant advancement in RNA detection technology that addresses specific limitations of these established methods while creating new opportunities for spatial transcriptomics.
RNAscope's unique value proposition lies in its ability to provide single-molecule sensitivity while maintaining spatial resolution at the single-cell level within intact tissue sections [2] [57]. This technical achievement bridges a critical gap between quantitative PCR's sensitivity and IHC's morphological context, creating a complementary role rather than a competitive one with existing methodologies. As systematic reviews have concluded, RNAscope serves as a "reliable and robust method that could complement gold standard techniques" rather than replace them [16].
Technical Mechanisms: The Foundation of RNAscope's Complementary Role
The Double-Z Probe Design: A Specificity Breakthrough
The fundamental innovation underlying RNAscope's performance is its proprietary double-Z probe design, which creates a mandatory two-step verification process for target detection [1] [57]. This system requires two separate "Z" probes to bind immediately adjacent to each other on the target RNA before signal amplification can proceed.
This dual-binding requirement dramatically reduces non-specific background signal that has plagued traditional ISH methods [1]. Each probe pair contains a target-specific sequence that hybridizes to the RNA of interest, connected by a linker sequence that provides the binding site for pre-amplifier molecules. The system employs approximately 20 different probe pairs targeting different regions of the same RNA molecule, ensuring consistent detection even if some target sites are obstructed [16].
Signal Amplification: Achieving Single-Molecule Sensitivity
Following successful probe binding, RNAscope employs a branched DNA (bDNA) amplification system that generates detectable signals without RNA replication [93]. This multi-stage amplification process enables visualization of individual RNA molecules as distinct dots under microscopy.
The sequential amplification process proceeds as follows:
- Pre-amplifier binding to the combined "Z" sequences of properly paired probes
- Amplifier molecules attaching to multiple sites on each pre-amplifier
- Labeled probes conjugating to amplifier molecules at a ratio of 20:1
This cascade results in up to 8,000-fold signal amplification for each target RNA molecule, enabling single-molecule detection without the background noise associated with traditional ISH amplification methods [16]. The amplification occurs without enzyme-based reactions, preserving tissue morphology and providing linear signal response that correlates with RNA expression levels.
Direct Technique Comparisons: Quantitative Performance Data
RNAscope vs. Traditional ISH and PCR Methods
Table 1: Performance Comparison Between RNAscope and Traditional Techniques
| Parameter | RNAscope | Traditional ISH | qRT-PCR | IHC |
|---|---|---|---|---|
| Sensitivity | Single-molecule detection [2] | Limited to highly expressed genes [16] | High (theoretical limit of 1 molecule) | Variable (depends on antibody affinity) |
| Specificity | High (100% reported) [16] | Moderate to low [16] | High | Variable (antibody cross-reactivity concerns) |
| Spatial Context | Preserved at single-cell level [49] | Preserved but limited by sensitivity | Lost (tissue homogenization) | Preserved at single-cell level |
| Target | RNA (≥300 nucleotides) [1] | RNA | RNA | Protein |
| Concordance with Gold Standards | 81.8-100% with qPCR/DNA ISH [16] | N/A | N/A | 58.7-95.3% with RNAscope [16] |
| Multiplexing Capability | Yes (multiple channels) [16] [49] | Limited | Limited (requires multiple reactions) | Limited (2-3 targets typically) |
The systematic review by PMC analyzing 27 studies demonstrated that RNAscope shows high concordance with qPCR and DNA ISH methods (81.8-100%), confirming its reliability for RNA detection [16]. However, the lower concordance with IHC (58.7-95.3%) highlights the fundamental differences between RNA and protein detection, with each method providing biologically complementary information about gene expression states.
RNAscope vs. Alternative ISH Technologies
Table 2: Comparison of Advanced RNA Detection Platforms
| Feature | RNAscope | HCR (Hybridization Chain Reaction) | BaseScope |
|---|---|---|---|
| Optimal Target Length | ≥300 nucleotides [1] | Varies | 50-300 nucleotides [1] |
| Probe Design | 20-25 base Z-probes [93] | Initiator/amplifier hairpin probes [93] | 1-6 ZZ probe pairs [1] |
| Signal Amplification | bDNA system [93] | Hybridization chain reaction [93] | bDNA system |
| Background Noise | Minimal due to double-Z specificity [1] | Moderate (background signal concerns) [93] | Minimal |
| Commercial Availability | Extensive catalog (>30,000 probes) [35] | Emerging commercial options | Specialized targets |
| Best Applications | mRNA, long non-coding RNA [1] | Flexible experimental designs | Short targets, splice variants, degraded RNA [1] |
When compared with HCR technology, RNAscope provides superior signal-to-noise ratio and more consistent results across different tissue types, particularly in challenging FFPE samples [93]. However, HCR may offer advantages in cost and probe design flexibility for researchers with specific custom requirements.
Experimental Applications: Methodologies and Implementation
Standardized RNAscope Workflow Protocol
The RNAscope procedure follows a structured workflow that can be implemented manually or automated, with specific quality control checkpoints essential for reliable results.
Sample Preparation Requirements
- Tissue Types: Compatible with FFPE (most common), fresh frozen, or fixed cells [16]
- Fixation: 10% neutral-buffered formalin for 16-32 hours recommended [57]
- Sectioning: 5μm thickness mounted on Superfrost Plus slides [6]
Critical Pretreatment Optimization
The systematic review emphasized that proper sample qualification using control probes is essential before target gene evaluation [16]. The standard protocol includes:
- Deparaffinization with xylene and ethanol series
- Antigen retrieval using boiling conditions (15 minutes at 95-100°C)
- Protease digestion (15-30 minutes at 40°C) for tissue permeabilization
Different tissue types require specific pretreatment optimization. For example, neural tissues (brain, spinal cord) typically require milder conditions (shorter protease treatment), while dense connective tissues may need extended retrieval [57].
Hybridization and Amplification
- Probe hybridization: Incubate target-specific probes for 2 hours at 40°C [57]
- Signal amplification: Sequential application of AMP1-AMP6 reagents (30 minutes each at 40°C) [57]
- Signal detection: Chromogenic (DAB/Red) or fluorescent detection compatible with standard microscopy
Essential Research Reagent Solutions
Table 3: Key Reagents for RNAscope Implementation
| Reagent/Category | Function | Examples & Specifications |
|---|---|---|
| Control Probes | Assay validation and sample qualification | PPIB (moderate expression), POLR2A (low expression), UBC (high expression), dapB (negative control) [16] [57] |
| Target Probes | Specific detection of RNA targets | >30,000 catalog probes; custom design in 2 weeks [49] [35] |
| Detection Kits | Signal generation and amplification | RNAscope 2.5 HD Brown/RED kits, multiplex fluorescent kits [57] |
| Pretreatment Reagents | Tissue preparation and permeabilization | Hydrogen peroxide block, epitope retrieval buffers, proteases [6] |
| Automation Systems | Standardized high-throughput processing | Leica BOND RX, Ventana DISCOVERY XT/ULTRA [57] [35] |
| Analysis Software | Signal quantification and interpretation | HALO, QuPath, Aperio algorithms [16] [57] |
Quality Control and Scoring Standards
Implementation of rigorous quality control measures is essential for reliable RNAscope results. The technology incorporates built-in control systems:
- Positive controls: Housekeeping genes (PPIB, POLR2A, UBC) validate RNA integrity [16]
- Negative controls: Bacterial dapB gene confirms absence of background staining [16]
- Scoring system: Semi-quantitative evaluation based on dots per cell [57]
The established scoring guidelines categorize results as:
- Score 0: No staining or <1 dot/10 cells
- Score 1: 1-3 dots/cell
- Score 2: 4-9 dots/cell, few clusters
- Score 3: 10-15 dots/cell, <10% clusters
- Score 4: >15 dots/cell, >10% clusters [57]
Strategic Implementation: When to Deploy RNAscope as a Complementary Technique
Ideal Applications for RNAscope
RNAscope provides maximum value when applied to specific research scenarios where its unique capabilities address critical questions:
Validation of PCR Findings with Spatial Context
Resolution of IHC Ambiguities
Cellular Heterogeneity Investigation
- When analyzing complex tissues with multiple cell types where bulk RNA analysis masks important differences [16]
- Example: Distinguishing tumor cell subtypes within the microenvironment
Low Abundance Target Detection
Multiplexed RNA Profiling
Integration with Established Workflows
The most effective implementation of RNAscope occurs when it's strategically positioned within a comprehensive experimental plan:
Hypothesis Generation Phase → RNAscope Spatial Validation → Mechanistic Investigation
- NGS/qPCR identifies candidate biomarkers → RNAscope localizes expression within tissue architecture → Functional studies explore biological significance
This integrated approach was exemplified in a veterinary cancer study where RNAscope provided complementary data to IHC for c-KIT evaluation in mast cell tumors, revealing statistically significant correlations with histological grade that protein detection alone failed to establish [94].
RNAscope occupies a distinctive and valuable position within the molecular biology toolkit, serving as a bridge between established techniques rather than a replacement for them. Its ability to provide spatial resolution of RNA expression at single-molecule sensitivity addresses a critical gap between PCR's quantitative power and IHC's morphological context. The experimental data demonstrates strong concordance with PCR methods while revealing important biological disparities with IHC that reflect the complex relationship between RNA and protein expression.
For researchers and drug development professionals, strategic implementation of RNAscope alongside existing methods enables more comprehensive biological understanding. By validating bulk RNA findings with spatial context, resolving ambiguous protein localization patterns, and characterizing cellular heterogeneity, RNAscope complements traditional techniques to create a more complete picture of gene expression dynamics. As with any technology, appropriate application based on specific research questions and sample considerations remains essential for maximizing its value in the constantly evolving landscape of molecular analysis.
Conclusion
The advent of RNAscope technology represents a paradigm shift in RNA in situ hybridization, moving the field from the detection of highly expressed genes to the precise visualization of individual RNA molecules within an intact morphological context. Its superior sensitivity and specificity, enabled by a unique probe design and amplification system, address the long-standing limitations of traditional ISH. This capability is not merely incremental; it is foundational, enabling researchers to validate high-throughput transcriptomic discoveries, unravel cellular heterogeneity in cancer and complex tissues, and accurately detect low-abundance viral RNAs. As validation in clinical settings grows and digital quantification methods become more sophisticated, RNAscope is poised to accelerate the transition of RNA biomarkers from research curiosities to robust tools for diagnostic pathology and targeted therapy development, ultimately paving the way for more personalized medicine approaches.
References
- 1. New-ISH on the Block: Introduction to RNAscope® [https://bitesizebio.com/40621/new-ish-on-the-block-introduction-to-rnascope/]
- 2. RNAscope: A Novel in Situ RNA Analysis Platform for ... [https://www.sciencedirect.com/science/article/pii/S1525157811002571]
- 3. RNAscope, In Situ Hybridization Technique | ACD [https://acdbio.com/referenceguide]
- 4. RNAscope Enables Spatial Genomic ISH ... [https://acdbio.com/science/how-it-works]
- 5. Deep Learning Segmentation of Chromogenic Dye ... [https://link.springer.com/article/10.1007/s10278-024-01301-9]
- 6. RNAscope Troubleshooting Guide and FAQ [https://acdbio.com/technical-support/solutions/faq]
- 7. Intronic RNAscope probes enable precise identification of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11977257/]
- 8. QuantISH: RNA in situ hybridization image analysis framework ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9249626/]
- 9. RNAscope: A Novel in Situ RNA Analysis Platform for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3338343/]
- 10. The optimisation and application of a novel BaseScope ... [https://www.sciencedirect.com/science/article/pii/S0166093425001302]
- 11. RNAscope Assay Technical Support and FAQ [https://acdbio.com/technical-support/getting-started]
- 12. RNAscope Workflow, In Situ Hybridization Steps | ACD [https://acdbio.com/ebook/introduction/rnascope-workflow]
- 13. Combining RNAscope and immunohistochemistry to ... [https://www.frontiersin.org/journals/molecular-neuroscience/articles/10.3389/fnmol.2023.1225847/full]
- 14. RNA sequencing and immunohistochemistry jointly ... [https://www.nature.com/articles/s41598-025-12780-3]
- 15. RNAscope Assay Workflow | In Situ Hybridization, RNA-ISH [https://acdbio.com/support-solution-category/rnascope-assay-workflow]
- 16. Evaluation of the Suitability of RNAscope as a Technique ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8710359/]
- 17. RNAscope in situ hybridization (ISH) FAQs [https://www.bio-techne.com/reagents/rnascope-ish-technology/getting-started/faqs]
- 18. RNAscope Probes [https://www.bio-techne.com/reagents/rnascope-ish-technology/probes]
- 19. bio-techne.com/reagents/ rnascope - ish -technology [https://www.bio-techne.com/reagents/rnascope-ish-technology]
- 20. A Method for Combining RNAscope In Situ Hybridization... | PLOS One [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0120120]
- 21. Reference Standards | In Situ Hybridization, RNA-ISH [https://acdbio.com/science/applications/research-solutions/reference-standards]
- 22. In Situ Hybridization - Histology Research Core [https://histologyresearchcorefacility.web.unc.edu/rna-scope/]
- 23. Intronic RNAscope probes enable precise identification of ... [https://www.nature.com/articles/s42003-025-08012-z]
- 24. See More in Vivid Color! Introducing the Newly Enhanced ... [https://info.acdbio.com/webinar-ACD-Vivid-od.html]
- 25. 2025 Mar 25 - Visualize and Quantify Oligonucleotide ... [https://acdbio.com/2025-mar-25-visualize-and-quantify-oligonucleotide-therapy-delivery-spatial-biodistribution-and]
- 26. RNAscope common questions, troubleshooting, and tips ... [https://indicalab.com/news/rnascope-common-questions-troubleshooting-and-tips-and-tricks-2/]
- 27. Effect of formalin-fixation and paraffin-embedded tissue ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11185121/]
- 28. RNAscope Multiplex FISH Signal Assessment in FFPE and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11755420/]
- 29. Control Slides and Control Probes for RNAscope [https://acdbio.com/control-slides-and-control-probes-rnascope]
- 30. 2025 Jan 14 - Unlock the role of spatial profiling in ... [https://acdbio.com/2025-jan-14-unlock-role-spatial-profiling-detecting-diverse-mrna-markers-and-short-targets]
- 31. Mastering Spatial RNA Detection: High Specificity & ... [https://news.ki.se/calendar/histocore-mastering-spatial-rna-detection-high-specificity-sensitivity-with-rnascope-ish-on-leica-bond-rx-by-acdbio]
- 32. RNAscope ISH [https://www.bio-techne.com/reagents/rnascope-ish-technology/rnascope]
- 33. Mapping human tissues with highly multiplexed RNA in situ ... [https://www.nature.com/articles/s41467-024-46437-y]
- 34. Bio-Techne Launches Expanded Menu of RNAscope ... [https://acdbio.com/bio-techne-launches-expanded-menu-rnascope-probes-human-and-mouse-transcriptome-advance-spatial]
- 35. RNAscope CE-IVD Detection | In Situ Hybridization, RNA-ISH [https://acdbio.com/rnascope-ce-ivd-detection]
- 36. Technical innovations of signal amplification in ... [https://pubmed.ncbi.nlm.nih.gov/40578770/]
- 37. RNAscope Multiplex Fluorescent V2 Assay | ACD [https://acdbio.com/rnascope-multiplex-fluorescent-v2-assay]
- 38. RNAscope™ Fluorescent Multiplex Assays by ACD [https://acdbio.com/rnascope-fluorescent-multiplex-assays]
- 39. Detection and Quantification of Multiple RNA Sequences ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7476812/]
- 40. Multiplexed RNA imaging and in situ profiling in living cells - Chemical Science (RSC Publishing) DOI:10.1039/D5SC06220A [https://pubs.rsc.org/en/content/articlehtml/2025/sc/d5sc06220a]
- 41. RNAscope™ Multiplex Fluorescent Assays | Bio-Techne [https://www.bio-techne.com/reagents/rnascope-ish-technology/rnascope/fluorescent-assays/multiplex]
- 42. NGS Validation with RNAscope ISH [https://acdbio.com/science/applications/research-solutions/ngsht-transcriptomic-validation-rna-seq]
- 43. RNAscope® technology | In Situ Hybridization, RNA-ISH [https://acdbio.com/dataanalysisguide]
- 44. A Guide for RNAscope™ Data Analysis [https://www.bio-techne.com/reagents/rnascope-ish-technology/rnascope/guide-rnascope-data-analysis]
- 45. Automated Quantitative RNA in Situ Hybridization for ... [https://www.sciencedirect.com/science/article/pii/S1525157812003157]
- 46. 2022 Jul 26 - Visualization and Analysis of RNAscope™ ... [https://acdbio.com/2022-jul-26-visualization-and-analysis-rnascope%E2%84%A2-results-using-qupath]
- 47. RNAscope Assay Visualization and Interpretation [https://acdbio.com/support-solution-category/rnascope-assay-visualization-and-interpretation]
- 48. 2019 Sept 24 - Validating single-cell RNA-seq data using ... [https://acdbio.com/2019-sept-24-validating-single-cell-rna-seq-data-using-single-cell-western-blotting-and-rnascope]
- 49. Viral RNA Detection using RNAscope® ISH | ACD [https://acdbio.com/science/applications/research-areas/infectious-diseases]
- 50. Validation of a DKK1 RNAscope chromogenic in situ ... [https://www.nature.com/articles/s41598-021-89060-3]
- 51. RNAscope Troubleshooting Guide and FAQ [https://acdbio.com/technical-support/solutions]
- 52. Regular article Comparative Analysis of Gene Expression ... [https://www.sciencedirect.com/science/article/pii/S1525157824001636]
- 53. Combining RNAscope and immunohistochemistry to visualize ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10470653/]
- 54. Best of 2024: Overview of insightful publications in diverse ... [https://acdbio.com/2025-feb-11-best-2024-overview-insightful-publications-diverse-research-areas-using-rnascope]
- 55. RNAscope ISH Troubleshooting | Bio-Techne [https://www.bio-techne.com/resources/protocols-troubleshooting/rnascope-ish-troubleshooting]
- 56. RNAscope in situ hybridization confirms mRNA integrity ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5706804/]
- 57. RNAscope ISH Reference Guide | Bio-Techne [https://www.bio-techne.com/reagents/rnascope-ish-technology/rnascope/rnascope-ish-reference-guide]
- 58. Using Single Molecule mRNA Fluorescent in Situ ... [https://www.nature.com/articles/s41598-018-26345-0]
- 59. Comparing non-machine learning vs. ... [https://www.nature.com/articles/s41598-025-08778-6]
- 60. Comparison of manual and automated digital image ... [https://pubmed.ncbi.nlm.nih.gov/35146728/]
- 61. Digital image analysis outperforms manual biomarker ... [https://www.sciencedirect.com/science/article/pii/S0893395222021123]
- 62. Quantitative Analysis of Gene Expression in RNAscope ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9901451/]
- 63. Digital pathology in tau research: A comparison of QuPath ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12277702/]
- 64. HALO | Quantitative Image Analysis for Pathology [https://indicalab.com/halo/]
- 65. QuPath vs Halo vs Visiopharm - Usage & Issues [https://forum.image.sc/t/qupath-vs-halo-vs-visiopharm/45310]
- 66. Comparison of QuPath and HALO platforms for analysis ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12139998/]
- 67. Catalog and Made-to-Order Probes from ACD [https://acdbio.com/all-about-probes]
- 68. Protocol for multiplexed RNAscope-based imaging of ... [https://www.sciencedirect.com/science/article/pii/S2666166725000218]
- 69. QuantISH: RNA in situ hybridization image analysis ... [https://www.nature.com/articles/s41374-022-00743-5]
- 70. Bio-Techne Expands RNAscope® Assay Services Portfolio ... [https://investors.bio-techne.com/press-releases/detail/97/bio-techne-expands-rnascope-assay-services-portfolio-to]
- 71. Protocol to validate custom-designed BaseScope probes ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12615734/]
- 72. RNA Hybridization, Multiplex Assays Central to Bio- ... [https://acdbio.com/rna-hybridization-multiplex-assays-central-bio-technes-growth-plans]
- 73. RNAscope compatibility with image analysis platforms for ... [https://www.sciencedirect.com/science/article/pii/S0065128121000878]
- 74. A Comparative Study with 10% Neutral Buffered Formalin [https://pmc.ncbi.nlm.nih.gov/articles/PMC12154963/]
- 75. Fresh Frozen RNAScope Protocol [https://www.protocols.io/view/fresh-frozen-rnascope-protocol-hfnab3maf.html]
- 76. RNAscope Webinars, Videos, Presentations | Learn More [https://acdbio.com/technical-support/learn-more]
- 77. A novel protein-preserving passive tissue clearing ... [https://www.nature.com/articles/s12276-025-01550-w]
- 78. Her2 assessment using quantitative reverse transcriptase polymerase chain reaction reliably identifies Her2 overexpression without amplification in breast cancer cases | Journal of Translational Medicine | Full Text [https://translational-medicine.biomedcentral.com/articles/10.1186/s12967-017-1195-7]
- 79. Concordance of RT-qPCR with immunohistochemistry and its beneficial role in breast cancer subtyping - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10519502/]
- 80. What are the differences between IHC and ISH and how do ... [https://www.enzo.com/note/what-are-the-differences-between-ihc-and-ish-and-how-do-they-help-in-the-fight-against-cancer/]
- 81. A Comparison Between Immunohistochemistry and mRNA ... [https://pubmed.ncbi.nlm.nih.gov/39471829/?utm_source=FeedFetcher&utm_medium=rss&utm_campaign=None&utm_content=14GVrEVieJsBLt532l8lZYTQD6RldlwpJeNCFcmBZFUC9pyMlO&fc=None&ff=20241031134211&v=2.18.0.post9+e462414]
- 82. Antibody Validation & Challenges [https://acdbio.com/science/applications/research-solutions/antibody-validation-and-challenges]
- 83. Comparison of RNA In Situ Hybridization and ... [https://pubmed.ncbi.nlm.nih.gov/32826529/]
- 84. IHC & ISH [https://research.pathology.wisc.edu/ihc-ish/]
- 85. Recent Advances in High-sensitivity In Situ Hybridization and Costs and Benefits to Consider When Employing These Methods - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10323200/]
- 86. Robust and sensitive in situ RNA detection using Yn-situ [https://pmc.ncbi.nlm.nih.gov/articles/PMC9046451/]
- 87. One probe fits all: a highly customizable modular RNA in situ ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12148028/]
- 88. Tetrahedral DNA dendritic nanostructure-enhanced FISH for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12537822/]
- 89. 2025 Mar 18 - Accelerate RNA and Protein Biomarker ... [https://acdbio.com/2025-mar-18-accelerate-rna-and-protein-biomarker-development-new-rnascope%E2%84%A2-ish-protease-free-assays]
- 90. In-Situ Hybridization Market Size to Hit USD ... [https://www.precedenceresearch.com/in-situ-hybridization-market]
- 91. Biomarker Assay Development | In Situ Hybridization, RNA ... [https://acdbio.com/biomarker-assay-development]
- 92. Tissue Based Companion Diagnostics in a Regulated Space [https://flagshipbio.com/tissue-based-companion-diagnostics-in-a-regulated-space/]
- 93. Choosing your In Situ project type [https://nif.hms.harvard.edu/choosing-your-situ-project-type]
- 94. RNAScope in situ Hybridization as a Novel Technique for ... [https://www.frontiersin.org/journals/veterinary-science/articles/10.3389/fvets.2021.591961/full]